Таксономические рубрики
Добавление материалов в HTML-формате на страницы Таксономических рубрик осуществляется под контролем Президента Общества. Это - авторские статьи, которые написаны специально для этого сайта и впервые размещены именно на нем, интеллектуальная собственность авторов. Поэтому тексты защищены от копирования и редактирования другими членами Общества. А вот фотографии, в основном, не наши. Они общедоступные, из интернета. Нам очень хотелось бы указать имя автора каждой фотографии. Ведь это его творческий продукт. Увы, это в большинстве случаев невозможно из-за многократного копирования-перекопирования. Но и просто красть фотографии с чужих сайтов нам мешает совесть. Вышли из положения так: возле каждого изображения есть активная ссылка на источник.
Если кто-то из Членов Общества хочет опубликовать главную статью какой-то рубрики (а мы в этом чрезвычайно заинтересованы), присылайте ее Президенту по электронной почте. Сохранение Вашего авторства и бережное отношение к этим материалам гарантируется. Все Члены Общества имеют право оставлять комментарии к центральной статье любой рубрики. Особенно поощряются указания на ошибки и неточности.
Подготовка главной статьи для каждой рубрики – это огромная и очень сложная работа. Поэтому не удивляйтесь, что многие рубрики долго будут оставаться без главной статьи. Мы начали наполнение сайта именно с таксономических рубрик. Причем, шли от экзотических таксонов к нашим и продвинулись уже довольно далеко. К настояему времени закончен первый этап этой работы: заполнены все таксономические рубрики, кроме семейства сосновые (Pinaceae). В настоящее время идет работа над этим самым важным для нас семейством. Надеемся, что имеющиеся пока пробелы будут отчасти восполнены добавлением других материалов в каждую рубрику: галерей, файлов, ссылок. Это могут делать все пользователи от Активного члена и выше: самостоятельно и без никаких ограничений.
Хвойные - это крупнейшая наиболее широко распространенная группа голосеменных. Кроме хвойных, к голосеменным относятся еще 3 современных группы (по современной таксономии - подклассы). Это саговниковые (Cycadidae), гинкговые (Ginkgoidae) и гнетовые (Gnetopidae). Общество у нас хвойное, но мы все-таки сочли целесообразным обстоятельно рассказать о голосеменных в целом, а также включить в таксономические рубрики эти три подкласса. Они не имеют большого значения для России и ее декоративного садоводства. Поэтому представлены на нашем сайте одной страницей каждый. Весь остальной огромный объем информации относится к подклассу хвойные (Pinidae).
Подклассы делятся на порядки, порядки на семейства. Например, у покрытосеменных сотни семейств объединены в десятки порядков. У хвойных число порядков, в основном, совпадает с числом семейств. Поэтому в нашей рубрикации порядки пропущены, а подкласс хвойные делится сразу на семейства.
Каждое из тех семейств, которые не имеют большого значения для России – араукариевые (Araucariaceae), головчатотисовые (Cepholotaxaceae), подокарповые (Podocarpaceae) и сциадопитисовые (Sciadopitiaceae) – представлено у нас единственной страницей. В остальных трех семействах, важных для России, выделены один (тисовые, Taxaceae) или несколько (кипарисовые, Cupressaceae; сосновые, Pinaceae) родов.
Этот же принцип использован и для родов. Малозначимые для России (например, цедрус, Cedrus) или монотипые (содержащие единственный вид, например, микробиота, Microbiota) представлены единственной страницей. Важные роды подразделены на несколько или много видовых страниц. При этом важные виды поименованы отдельно, а для не особенно важных есть рубрика «Другие».
Если вы сейчас кликнете кнопку «Подробнее…», то попадете на страницу с подробной характеристикой голосеменных. Там довольно «многа букав». Просим за это прощения у читателя. В свое оправдание скажем, что это не просто буквы. Там собрана и систематизирована информация, которой на русском языке больше нет нигде. Собрана она по крупицам, главным образом, из англоязычных источников. Мы по возможности старались адаптировать ее для людей без специальной научной подготовки.
Приятного вам плавания по безбрежному морю хвойной и «околохвойной» информации!
Современная система голосеменных
Начнем с того, есть ли вообще голосеменные «как класс». В течение длительного времени они считались четкой естественной (т.е. имеющей одного общего предка) группой, а именно, отделом. Вторым таким же отделом считались цветковые. Это представление и система голосеменных из 6 классов (2 вымерших, 4 современных) существовало на всем протяжении 20-го века и казалось нам таким же устойчивым, как климат и руководящая роль КПСС. Причем, доминировало оно не только у нас, но и по всему миру.
 В СССР его поддерживал очень уважаемый, в том числе, на мировом уровне, человек – директор Ботанического института АН, академик Армен Леонович Тахтаджян (оно лучше всего обосновано в его работе «Высшие таксоны сосудистых растений, исключая цветковые». Проблемы палеоботаники. Л., 1986, с. 135-142). Армен Леонович большое внимание уделял популяризации ботанических знаний, в частности, лично руководил изданием последних томов знаменитой многотомной «Жизни растений» (1974-1982), посвященных высшим растениям. Естественно, в 4-м томе этого издания, где речь идет про голосеменные, они описаны в системе из упомянутых выше 6 классов. Ссылка на источник фотографии.
В СССР его поддерживал очень уважаемый, в том числе, на мировом уровне, человек – директор Ботанического института АН, академик Армен Леонович Тахтаджян (оно лучше всего обосновано в его работе «Высшие таксоны сосудистых растений, исключая цветковые». Проблемы палеоботаники. Л., 1986, с. 135-142). Армен Леонович большое внимание уделял популяризации ботанических знаний, в частности, лично руководил изданием последних томов знаменитой многотомной «Жизни растений» (1974-1982), посвященных высшим растениям. Естественно, в 4-м томе этого издания, где речь идет про голосеменные, они описаны в системе из упомянутых выше 6 классов. Ссылка на источник фотографии.
Всегда были противники этой точки зрения, которые сомневались в монофилетическом происхождении голосеменных. Эта гипотеза к настоящему времени получила веские доказательства и стала почти общепризнанной теорией. Радикальные изменения произошли из-за молекулярной революции в филогенетике и, соответственно, систематике, которая началась на рубеже прошлого и нынешнего веков. Если раньше о родстве разных групп судили по морфологическим признакам, которые каждый исследователь выбирал и толковал по своему разумению, то теперь о родстве стали судить непосредственно по структуре ДНК, что резко сократило возможности субъективных оценок.
По результатам очень многих исследований примерно к 2010 г. сформировалась вот такая система высших растений (Chase, M.W. & Reveal, J.L. (2009) A phylogenetic classification of the land plants to accompany APG III. Botanical Journal of the Linnean Society 161: 122–127):
Class Equisetopsida
Subclass 1. Anthocerotidae
Subclass 2. Bryidae
Subclass 3. Marchantiidae
Subclass 4. Lycopodiidae
Subclass 5. Equisetidae
Subclass 6. Marattiidae
Subclass 7. Ophioglossidae
Subclass 8. Polypodiidae
Subclass 9. Psilotidae
Subclass 10. Ginkgoidae - гинкговые
Subclass 11. Cycadidae - саговниковые
Subclass 12. Pinidae - хвойные
Subclass 13. Gnetidae - гнетовые
Subclass 14. Magnoliidae - покрытосеменные
Как видите, по этой новой системе все без исключения сосудистые растения (если говорить по простому, плауны + хвощи + папоротники + голосеменные + покрытосеменные) относятся к одному классу Equisetopsida (Хвощевые). Нет, это не опечатка. Почему именно хвощевые, объяснить можно: так велит кодекс ботанической номенклатуры, это связано с приоритетностью названий. Но в тонкости вникать не будем. Оставим название на совести ботаников. Для нас важно, что этот класс – монофилетический: имеет одного общего предка. Обратите внимание, что это даже не отдел, а всего лишь класс, который сформировали из 6 (!) прежних отделов. Тут отразилась общая тенденция современной филогенетики - зреть в корень: главные «развилки» древа растений, породившие отделы, произошли задолго до появления высших растений.
Внутри класса Equisetopsida 14 подклассов. Значит, все 14 ветвей отделялись от основного ствола филогенетического древа по очереди и по одной, т.е. являются монофилетическими. Из них первые 9 подклассов относятся к высшим споровым: плауны, хвощи, папоротники. Почему их не 3, а 9, разбираться не будем, поскольку это не наша тема. 14-й подкласс Magnoliidae (покрытосеменные или цветковые) полностью совпадает с прежним отделом Magnoliophyta. А вот отдел Pinothyta (голосеменные) исчез без следа. Он отсутствует даже «как класс». Тем более, как отдел. На его месте появилось 4 подкласса (обозначены жирным шрифтом), прямо не связанных между собой и ничем не объединенных. Получается, настоящий раздел – о том, чего вообще нет в природе: о когда-то существовавшей в воображении ученых единой группе голосеменных растений.
Какое значение имеет этот, казалось бы, прискорбный факт? Очень большое: для тех исследователей, которые профессионально занимаются происхождением и родственными связями высших таксонов растительного мира. Для нас же - ни малейшего. Ведь все 4 привычных нам группы голосеменных – саговниковые, гинкговые, гнетовые и хвойные – ПОКА (почему пока, расскажем чуть позже) сохранились в прежнем объеме и даже под прежними названиями. Никуда не исчезли также их общие свойства, из-за которых их когда-то объединяли. Поэтому в настоящее время их продолжают объединять, правда, лишь в неформальную группу подклассов. Оно и понятно: ведь это 4 ветви, которые последовательно, одна за другой, отделились от общего ствола высших растений. А посему не будем драматизировать ситуацию. Спокойно рассмотрим структуру разнообразия в этой части филогенетического древа растительного мира.
Происхождение голосеменных, родственные связи
Начнем, ясное дело, с истории. Эволюция высших растений – большой и важный вопрос. Его изучают с самых разных сторон: от идентификации окаменелостей до тонких анализов ДНК. Поэтому, если набрать в поисковике три слова “land plant phylogeny” (филогения наземных растений), то на пользователя сразу вываливается такой объем информации, что даже выбраться из под него, не говоря уже о том, чтобы разобраться в нем, оказывается совершенно невозможно без специальной подготовки и запредельной мотивации. Да и зачем оно нам? Ведь это не совсем наша тема. Поэтому ограничимся лишь одной скромной картинкой, самой простой из всех возможных.

Филогения зеленых растений. На ней хорошо показан основной принцип их эволюции: от основного ствола последовательно отчленяются всё новые, более продвинутые ветви. Зеленые водоросли (green algae) появились на нашей планете 1,5 млрд. лет назад. Чтобы выйти на сушу, растениям потребовалось не много и не мало, а ровно один миллиард лет. От общего ствола наземных растений первыми 500 млн. лет назад отделились мохообразные (mosses). Еще через 80 млн. лет обособились первые сосудистые растения. Далее схема сильно упрощена. Сосудистые растения представлены всего четырьмя ветвями: планувидными (lycophytes), папоротниковидными (ferns), голосеменными (gymnosperms) и покрытосеменными (monocots + dicots = flowering plants). На самом деле, как мы уже писали выше, таких самостоятельных ветвей было не 4, а 14. Объект нашего интереса, 4 ветви, условно относящиеся к голосеменным, начали отчленяться от основного ствола 350 млн. лет назад. После этого потребовалось еще 200 млн. лет, чтобы вершина филогенетического древа начала активно дифференцироваться как подкласс покрытосеменные. Ссылка на источник рисунка.
Чем голосеменные отличаются от относительно ранних ответвлений филогенетического древа и от последнего достижения природы – покрытосеменных? До появления первых голосеменных зеленые растения провели на суше уже довольно много времени. Выход на сушу был для растения явным выходом из «зоны комфорта»: он сильно способствовал их прогрессивному развитию, в первую очередь, дифференциации ВЕГЕТАТИВНОГО тела. Новая среда обитания состояла их двух принципиально разных во всех отношениях половинок: почвенной и воздушной. Для каждой из них сформировались соответствующие органы, корень и лист, а также соединяющий их стебель.
Дифференциация органов происходила параллельно с дифференциацией тканей. В не родной для растений воздушной среде им надо было, с одной стороны, не дать себе засохнуть, с другой, обеспечить газам полный доступ внутрь листа для целей фотосинтеза и дыхания. Так появилась покровная ткань, эпидерма, со сложно организованными клапанами - устьицами. Однако устьица пропускали также и воду: в виде пара (транспирация). Транспирация использовалась для охлаждения листа в случае необходимости. Вода – это и среда, и материал для фотосинтеза. Поступала же она вместе с минеральными элементами теперь только из корней. Чтобы корни эффективно выполняли эту свою функцию, их надо было хорошо кормить углеводами. Появилась необходимость «гонять» воду с разными растворенными в ней веществами от корней к листьям и обратно.
Ответом на эту необходимость стало появление и эволюция проводящих тканей. Самая сложная задача - поднимать воду вверх, от корней к листьям, против вектора гравитации. По живым клеткам она быстро и эффективно подниматься не может. Пришлось часть клеток для этой цели умертвить. Мертвые клетки с обычными тонкими целлюлозными стенками не могут располагаться среди живых потому, что не могут сопротивляться их давлению. Значит, перед умерщвлением надо сначала обеспечить клеточным стенкам достаточную механическую прочность. Растения решили эту проблему через их утолщение и лигнификацию (одревеснение).
Последнее удачно совпало с необходимостью обеспечить органам общую механическую устойчивость в обеих агрессивных средах: и в жесткой минеральной и в динамичной (ветер) воздушной. Так проводящие ткани стали выполнять дополнительную, механическую функцию. Появились также и специализированные механические ткани.
Необходимость организации и гармонизации сложнейшей системы органов и тканей, которая характерна для наземных растений, вызвала появление адекватного по сложности и эффективности механизма саморегуляции морфогенеза. В нем решающая роль принадлежит специализированным образовательным тканям (меристемам): верхушечным (отвечающим за новообразование органов и рост в длину) и боковым (отвечающим за рост в толщину).
Всё это отлично выполняло свои функции, гармонично сочеталось и взаимодействовало. С одной лишь оговоркой: в теплом и влажном климате, т.е. в новой «зоне комфорта». Стремление выйти из нее вызвало новую мощную «реформу», на этот раз в сфере ПОЛОВОЙ РЕПРОДУКЦИИ.
 Лес каменоугольного периода. Последний назван так потому, что из органического вещества таких лесов образовались современные залежи каменного угля. Отсюда понятно, насколько мощными были эти леса из древовидных плаунов, хвощей и папоротников. Обратите внимание на обилие воды в лесу. Это не случайно. Ссылка на источник рисунка.
Лес каменоугольного периода. Последний назван так потому, что из органического вещества таких лесов образовались современные залежи каменного угля. Отсюда понятно, насколько мощными были эти леса из древовидных плаунов, хвощей и папоротников. Обратите внимание на обилие воды в лесу. Это не случайно. Ссылка на источник рисунка.
Жесткая привязка плаунов, хвощей и папоротников к теплу и сырости определялась, главным образом, их репродуктивной системой. Не зря ведь названия трех основных ступеней развития наземных растений характеризуют способ их размножения: высшие споровые, голосеменные, покрытосеменные. Действительно, эволюция их репродуктивной системы была «локомотивом» общего прогресса. Тут мы вступаем в тему так называемых жизненных циклов, самую муторную область эволюционной ботаники, которую лютой ненавистью ненавидят многие поколения студентов. Простые любители ботаники, изредка открывающие умные книги по систематике растений, обычно пролистывают, не глядя, эти страницы, чтобы добраться поскорее до более приятных и понятных. Ну а нам деваться некуда: придется разбираться. Иначе мы просто не сможем продвинуться дальше. В конце концов, ничего сильно страшного тут нет. Это всего лишь информация из школьного учебника ботаники.
 Итак, жизненный цикл высших споровых. Рассмотрим его на примере одного из наших папоротников. То, что мы обычно видим в лесу, многолетнее травянистое растение с розеткой крупных перистых листьев, в терминологии жизненного цикла представляет собой спорофит. Называется так потому, что дает споры. Они формируются в спорангиях, расположенных на нижней стороне листа. Спорофит диплоидный: в его клетках двойной набор хромосом (2n). Споры гаплоидные (n). Поэтому образованию спор предшествует мейоз – редукционное деление, сокращающее (редуцирующее) число хромосом вдвое. Споры мелкие, легкие, пылевидные. После созревания они разносятся ветром.
Итак, жизненный цикл высших споровых. Рассмотрим его на примере одного из наших папоротников. То, что мы обычно видим в лесу, многолетнее травянистое растение с розеткой крупных перистых листьев, в терминологии жизненного цикла представляет собой спорофит. Называется так потому, что дает споры. Они формируются в спорангиях, расположенных на нижней стороне листа. Спорофит диплоидный: в его клетках двойной набор хромосом (2n). Споры гаплоидные (n). Поэтому образованию спор предшествует мейоз – редукционное деление, сокращающее (редуцирующее) число хромосом вдвое. Споры мелкие, легкие, пылевидные. После созревания они разносятся ветром.
Попадая на почву, в относительно теплую и влажную среду, спора прорастает. Из нее формируется так называемый заросток, более или менее параллельная почве зеленая пластинка размером до 1 см с корнеподобными выростами (ризоидами). Заросток самодостаточен: он фотосинтезирует, поглощает воду из почвы. Правда, живет недолго: неделю-другую-третью. В терминологии жизненного цикла заросток представляет собой гаметофит: будучи гаплоидным, он производит такие же гаплоидные половые клетки - гаметы. Они, естественно, двух типов: мужские (сперматозоиды) в мужских гаметангиях (антеридиях), женские (яйцеклетки) в женских гаметангиях (архегониях).
На рисунке мы видим, что гаметангии располагаются как будто бы на верхней стороне заростка. Нет, художник перевернул его на 180 градусов. На самом деле, гаметангии располагаются строго на нижней стороне: там, где выше влажность. Созревая, яйцеклетки остаются в архегонии. Сперматозоиды же выходят во внешнюю среду и призваны совершить подвиг: героически доплыть до яйцеклетки по воде во время дождя. Некоторым это удается. Победитель оплодотворяет яйцеклетку. Получается диплоидная (n + n = 2n) зигота. Она делится, дифференцируется. Из нее формируется спорофит следующей генерации. Цикл замкнулся. Ссылка на источник рисунка.
Казалось бы, всё это отлично «придумано» природой. Нет, не отлично. Основной смысл полового процесса такой: в нем участвуют заведомо разные папа и мама, которые по этой причине дают здоровое, а главное, РАЗНООБРАЗНОЕ потомство, как материал для естественного отбора. Спрашивается, какой же смысл затевать половой процесс, если он обеспечивает лишь самооплодотворение: ведь вероятность того, что данную яйцеклетку оплодотворит не просто герой, а «супергерой», производный от соседнего заростка, стремится к нулю. Похоже, именно поэтому ТАКИЕ папоротники оказались тупиковой ветвью эволюции, а ее стволом на время стали другие папоротники: РАЗНОСПОРОВЫЕ.
Суть новации в том, что мужские гаметы образуются на разных гаметофитах (мужском и женском), которые вырастают из разных (соответственно, микро- и мега-) спор, которые формируются, соответственно, в разных (микро- и мега-) спорангиях. При таком раскладе вероятность самооплодотворения резко снижается чуть не до нуля: ведь микро и мегаспоры свободно распространяются по территории, а производные от них гаметофиты случайно перемешаны в пространстве. Тем самым достигается главная цель полового процесса: рекомбинация генов и признаков. И всё бы хорошо, да что-то нехорошо. Когда мужские и женские гаметы образуются рядом, под одним малюсеньким заростком (у равноспоровых папоротников), то сперматозоиды, собравшись с силами, успевают за период дождя в спринтерском темпе преодолеть вплавь несколько миллиметров, отделяющих их от яйцеклетки. Если мужские и женские гаметы образуются на разных заростках, пусть даже расположенных на расстоянии всего нескольких сантиметров друг от друга (а ведь и это маловероятно), то несчастным сперматозоидам не остается никаких шансов выполнить свою миссию. Именно поэтому среди современных папоротников разноспоровыми являются только 3 маленьких семейства с чисто водным образом жизни. В воде у сперматозоидов хотя бы нет дефицита времени: можно отдохнуть, собраться с силами и плыть дальше. Чтобы совместить разноспоровость (значит, и перекрестное оплодотворение) с жизнью на суше, растениям нужно было «придумать» что-то еще.
Они пошли по пути максимально возможной редукции гаметофитов с тем, чтобы не допускать их, слабых и беззащитных, до самостоятельной жизни во враждебной и непредсказуемой внешней среде. Чтобы они полностью находились под контролем и под защитой спорофитов, на их полном довольствии. А главное, чтобы оплодотворение максимально, желательно – полностью, было независимо от наличия капельно-жидкой воды. Эволюция растений в этих направлениях удивительным образом автоматически привела к появлению принципиального новообразования – семени как универсального органа размножения. Эти процессы были не быстрыми, заняли миллионы лет, прошли через много промежуточных этапов. Мы это всё пропустим, а рассмотрим сразу наполовину готовый продукт – жизненный цикл современных голосеменных. Пожалуйста, сосредоточьтесь и наберитесь терпения. В тонкие тонкости мы вникать не будем: обсудим лишь принцип.
 Типичный представитель голосеменных, сосна обыкновенная. Само дерево – это, естественно, диплоидный спорофит. Значит, он после редукционного деления (мейоза) генерирует гаплоидные споры. Разные, микро- и мега-, поскольку сосна, как и все семенные растения, относится к разноспоровым. Из микроспоры формируется мужской, из мегаспоры – женский гаметофит. Поэтому всю микро-линию у голосеменных часто называют мужской, а макро-линию – женской.
Типичный представитель голосеменных, сосна обыкновенная. Само дерево – это, естественно, диплоидный спорофит. Значит, он после редукционного деления (мейоза) генерирует гаплоидные споры. Разные, микро- и мега-, поскольку сосна, как и все семенные растения, относится к разноспоровым. Из микроспоры формируется мужской, из мегаспоры – женский гаметофит. Поэтому всю микро-линию у голосеменных часто называют мужской, а макро-линию – женской.
Микроспоры образуются в микроспорангиях, из которых состоит мужская шишка. Несколько упрощая ситуацию, можно сказать, что микроспоры – это пыльцевые зерна. На самом деле, то пыльцевое зерно, которое мы видим – это уже не совсем микроспора, потому, что в ней (прямо внутри) началось развитие мужского гаметофита.
Мегаспоры образуются в мегаспорангиях, которые формируются в женской шишке. У семенных растений мегаспорангий совсем не такой, как у споровых. Он имеет собственное название «нуцеллус» и окружен довольно жестким защитным покровом – интегументом. Интегумент + нуцеллус = семяпочка (семязачаток). Семяпочка есть важнейшее новообразование, отличающее семенные растения от высших споровых. В каждом мегаспорангии образуется одна-единственная функционирующая мегаспора. Прямо внутри мегаспорангия она многократно делится, формирует женский (мега-) гаметофит.
Таким образом, оба гаметофита даже и «не помышляют» о самостоятельном существовании: они живут за счет спорофита, развиваются внутри его тканей и органов. Развившись до некоторой стадии, пыльцевое зерно, (т.е. мужской гаметофит в оболочке микроспоры) покидает подсохшую мужскую шишку. Большая часть пыльцевых зерен, естественно, летят в никуда. Однако их настолько много, что часть всё-таки попадает на семяпочки. Последние имеют специальное отверстие для пыльцы: микропиле. В период лёта пыльцы оно заполнено специальной жидкостью. Это так называемая опылительная капля. Пыльца попадает не ее поверхность. Капля подсыхает и втягивает пыльцевые зерна внутрь семяпочки. Так мужской гаметофит попадает на поверхность женского гаметофита.
Мужской гаметофит у сосны сильно редуцирован. Путь от микроспоры до мужских гамет – это всего пять клеточных делений. Мужских гаметангиев (антеридиев), естественно, нет. Есть единственная антеридиальная клетка. Женский гаметофит, наоборот, довольно мощный, очень многоклеточный. На нем есть настоящие, «несколькоклеточные» архегонии. Обычно их два, в каждом созревает по одной яйцеклетке.
Мужской гаметофит, когда приходит время, образует одноклеточную пыльцевую трубку, которая внедряется в женский гаметофит и растет в сторону архегониев. Внутри, в концевой части, находятся спермии. Они так называются потому, что в отличие от сперматозоидов, неподвижны. Подвижность не требуется, т.к. кончик пыльцевой трубки доставляет их строго по адресу: до неподвижной яйцеклетки. Происходит оплодотворение, образуется зигота, она дифференцируется в зародыш. Женский гаметофит становится эндоспермом – питательной тканью семени. Она гаплоидная. Из покрова семяпочки – интегумента – образуется семенная кожура. Семя лежит голо на поверхности семенной чешуи, оно ничем не покрыто. Отсюда название «голосеменные». Семя созревает, покидает раскрывшуюся шишку, прорастает, из проростка формируется спорофит следующей генерации. Ссылка на источник рисунка.
Семя – несравненно более надежная единица расселения, чем спора. В семени находится дифференцированный, полностью готовый к прорастанию зародыш. А также запас питательных веществ и ферментов, необходимых для их использования, позволяющий зародышу прорасти, а проростку – начать самостоятельную жизнь. Всё это надежно защищено семенной кожурой.
Это если коротенько. На самом деле, всё сложнее, есть много важных и интересных подробностей. Мы к ним еще неоднократно вернемся в разных разделах. Здесь же сделаем некоторое обобщение. В части жизненного (репродуктивного) цикла голосеменные сделали гигантский шаг вперед по сравнению с высшими споровыми. Отметим 3 грандиозных преобразования. (1) Гаметофиты, особенно мужской, сильно редуцировались. Из самостоятельных зеленых растений они превратились (физиологически) в органы спорофита, тем самым оказались под его надежной защитой. Это резко повысило эффективность воспроизводства. (2) Репродуктивный цикл совершенно перестал быть зависимым от капельно-жидкой воды. Это позволило совместить разноспоровость с наземным образом жизни, тем самым резко ускорило темпы эволюции и резко расширило экологическую нишу. (3) Появилось семя с надежной оболочкой и запасом питательных веществ для зародыша как универсальный орган полового размножения.
 Все это в совокупности сильно способствовало расширению климатического и экологического ареала голосеменных, их процветанию и доминированию на протяжении почти всей мезозойской эры. На снимке представлена реконструкция «лес юрского периода». Плаунов, хвощей и папоротников всё ещё много. Но это уже, по большей части, травянистые растения. Среди древесных же абсолютно преобладают саговниковые, гинкговые и хвойные. Ссылка на источник рисунка.
Все это в совокупности сильно способствовало расширению климатического и экологического ареала голосеменных, их процветанию и доминированию на протяжении почти всей мезозойской эры. На снимке представлена реконструкция «лес юрского периода». Плаунов, хвощей и папоротников всё ещё много. Но это уже, по большей части, травянистые растения. Среди древесных же абсолютно преобладают саговниковые, гинкговые и хвойные. Ссылка на источник рисунка.
Увы, ничто не вечно под луной. Всё проходит. Прошел и период процветания голосеменных. Описанные выше достижения не спасли их от полного и безоговорочного поражения в борьбе за существование с покрытосеменными в конце мезозоя – начале кайнозоя. Почему так получилось? Чтобы уже раз и навсегда завершить эпопею с жизненными циклами, рассмотрим напоследок таковой у победителей и нынешних доминантов растительного покрова земли. Заодно выясним, почему они победили.
Размножение цветковых

У покрытосеменных в части жизненного цикла сохранились все достижения голосеменных + добавилось еще несколько новых: очень важных и полезных. Так, вместо шишки у них появился более совершенный орган половой репродукции - цветок. Отсюда их второе название: цветковые. В отличие от шишек, цветок в норме обоеполый. Казалось бы, это минус: должна повышаться вероятность нежелательного самооплодотворения. На самом деле, у цветковых есть очень эффективные средства, предотвращающие самооплодотворение. Поэтому обоеполый цветок совершенно не мешает им быстро эволюционировать.
Что нового и хорошего есть в цветке по сравнению с шишкой. Начнем с цветных лепестков, нектарников, запаха. Ничего похожего почти нет у голосеменных, потому что они опыляются по большей части ветром и поэтому вынуждены производить зазря огромную массу пыльцы. Цветковые опыляются, в основном, животными: прицельно. Кстати, тычинка (пыльник на длинной тонкой нити) как продукт эволюции простого микроспорофилла, по-видимому, тоже есть адаптация к опылению насекомыми. Последнее дает огромную экономию ресурсов: пыльцы требуется на несколько порядков меньше. Но главная новация в цветке - это, конечно, пестик с завязью и рыльцем. Завязь как раз и обеспечивает покрытосемянность: в ней, а также в образующемся из нее плоде, семяпочки и семена эффективно защищены на всем протяжении их развития вплоть до созревания. Рыльце также является важнейшим новообразованием (в истории даже были попытки назвать покрытосеменные рыльцевыми). Оно улавливает пыльцу: предназначено специально для этого.
Теперь о том, что невооруженным глазом не видно. Тут налицо два важных достижения: дальнейшая редукция гаметофитов и двойное оплодотворение. Как вы помните, у голосеменных путь от микроспоры до спермиев состоял всего из 5 клеточных делений. Неужели можно его еще сократить? Можно: в 2,5 раза! У покрытосеменных путь от микроспоры до спермиев – это всего 2 деления: (1) деление микроспоры на вегетативную клетку (из нее потом образуется пыльцевая трубка) и генеративную клетку, и (2) деление генеративной клетки на 2 спермия. Получается, что у покрытосеменных мужской гаметофит редуцирован до единственной клетки. Просто и ОЧЕНЬ эффективно.
Женский гаметофит голосеменных состоит из большого и неограниченного числа клеток, имеет архегонии, тоже слегка многоклеточные. У покрытосеменных ничего этого нет. Их женский гаметофит называется «зародышевый мешок». Его размер относительно стандартный: 0,003 мм. Это в десятки, а то и в сотни раз меньше, чем у голосеменных. Женский гаметофит цветковых состоит строго из 7 клеток: не больше и не меньше. Одна из них – яйцеклетка. Она ведет себя обыкновенно, как у всех: оплодотворяется спермием; из зиготы развивается зародыш.
На остальные 6 клеток зародышевого мешка приходится 7 ядер. Как так? В самом центре зародышевого мешка формируется огромная центральная клетка с двумя ядрами, которые вскоре сливаются, образуя одно диплоидное ядро. Зачем? Об этом чуть позже, а пока вернемся к спермиям. Их всегда образуется два. Один предназначен для оплодотворения яйцеклетки. А второй? Когда мы описывали жизненный цикл голосеменных, этот вопрос был несущественным. Второй спермий у голосеменных просто запасной. В абсолютном большинстве случаев он спокойно рассасывается, не найдя себе применения. Однако покрытосеменные не позволяют себе такой расточительности. У них без толку не пропадает ни единой клетки, тем более, половой. Второй спермий сливается с диплоидным ядром центральной клетки. Клетка становится триплоидной! Это есть второе оплодотворение. Из вновь образовавшейся клетки формируется триплоидный эндосперм – питательная ткань семени. Получается, что у цветковых эндосперм имеет совершенно другое происхождение по сравнению с голосеменными. И образуется он не заранее, а лишь тогда, когда уже точно известно, что оплодотворение яйцеклетки произошло. Этим достигается существенная экономия материальных ресурсов. Кроме того, похоже, что триплоидная пища (особенно по сравнению с гаплоидным эндоспермом голосеменных) есть «пища богов». Она более эффективно питает зародыш и проросток цветковых. Ссылка на источник рисунка.
 Продвинутый жизненный цикл – это не единственное преимущество цветковых над голосеменными. Много чего хорошего есть у них и в вегетативной сфере. Приведем только один пример. Их ксилема (древесина) в качестве проводящих элементов включает не только трахеиды, но и сосуды. В подробности пока вдаваться не будем. Посмотрим лишь на картинку. Это древесина клена. Узкие трубки – трахеиды, широкие – сосуды (трахеи). У голосеменных все трубки узкие, да еще и не сквозные, а с мелкими порами. Представьте, насколько тяжелей им качать воду вверх! Ссылка на источник рисунка.
Продвинутый жизненный цикл – это не единственное преимущество цветковых над голосеменными. Много чего хорошего есть у них и в вегетативной сфере. Приведем только один пример. Их ксилема (древесина) в качестве проводящих элементов включает не только трахеиды, но и сосуды. В подробности пока вдаваться не будем. Посмотрим лишь на картинку. Это древесина клена. Узкие трубки – трахеиды, широкие – сосуды (трахеи). У голосеменных все трубки узкие, да еще и не сквозные, а с мелкими порами. Представьте, насколько тяжелей им качать воду вверх! Ссылка на источник рисунка.
Лука Александрович у А.П.Чехова говорил Каштанке так: «Супротив человека ты всё равно, что плотник супротив столяра». Примерно то же самое можно сказать относительно голосеменных и цветковых. Последние не зря оказались победителями. Тем больше голосеменные нуждаются в нашей любви и дружбе. А посему продолжим и перейдем к их разнообразию.
Разнообразие современных голосеменных
Ботаники – народ дотошный. У них все ходы и виды (а также роды, семейства, порядки и т.д.) записаны в анналах. Не так давно вышла вот такая работа (Christenhusz, M. J., & Byng, J. W. (2016). The number of known plants species in the world and its annual increase. Phytotaxa, 261(3), 201–217), в которой проведен анализ наличного разнообразия сосудистых растений. Вот таблица, которую я составил по данным из этой публикации:

С тех пор информация, конечно, устарела: в этой же статье сказано, что в течение последних 25 лет число описанных наукой видов сосудистых растений ежегодно увеличивалось в среднем на 2 000. Тем не менее, из этой таблицы мы можем видеть, что голосеменные – это самая бедная видами, родами, семействами и порядками группа сосудистых растений. По всем этим показателям она уступает не только покрытосеменным (число видов в 274 раза меньше), но и высшим споровым (число видов в 11 раз меньше). Значительные различия между тремя группами наземных растений есть и по структуре разнообразия. Они показаны еще в одной таблице, составленной по результатам той же статьи:

Когда какая-то группа растений молода и находится на пике процветания, ее филогенетическое древо выглядит очень мощным, ветвистым и густым: в родах много видов, в семействах много родов, в порядках много семейств, в подклассах много порядков. У нас это, конечно, покрытосеменные. Структура разнообразия высших споровых весьма своеобразна. На их древность и, по сути, реликтовый характер указывает малое число порядков на подкласс, семейств на порядок, родов на семейство. Однако поразительная наполненность родов видами (в 2,5 раза выше, чем у покрытосеменных!) однозначно указывает, что у этой группы совсем недавно, примерно 5-10 млн. лет назад явно появилось «второе дыхание» и началось «вторая молодость». Голосеменные даже на их фоне выглядят бедно и бледно: у них налицо минимальное разнообразие на всех уровнях организации. Образованный ими фрагмент кроны общего филогенетического древа, в основном, усох. Лишь на некоторых очень далеко отстоящих друг от друга ветвях еще теплится жизнь.
 Проиллюстрируем эту мысль вот таким простым рисунком: распределение родов голосеменных по числу видов. Как видите, 41% родов – монотипые (состоят из единственного вида). Лишь три рода имеют около 100 видов или чуть больше: саговник (Cycas), подокарпус (Podocarpus) и сосна (Pinus). Источник рисунка - X.-Q. Wang, J.-H. Ran. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution. 75 (2014) 24–40.
Проиллюстрируем эту мысль вот таким простым рисунком: распределение родов голосеменных по числу видов. Как видите, 41% родов – монотипые (состоят из единственного вида). Лишь три рода имеют около 100 видов или чуть больше: саговник (Cycas), подокарпус (Podocarpus) и сосна (Pinus). Источник рисунка - X.-Q. Wang, J.-H. Ran. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution. 75 (2014) 24–40.
Почему так получилось? Казалось бы, мы уже ответили на этот вопрос: потому, что у цветковых оказались очень много полезных «инноваций». Всё верно. Но почему их оказалось так много, а у голосеменных инновации высокого уровня с некоторых пор перестали появляться. Видимо, потому, что у каждой более или менее перспективной филогенетической группы вначале её развития есть некий «звездный час», когда она молода и пластична, потентна и склонна к прогрессу. Такой период продолжается миллионы лет, но по геологическим меркам это не более, чем час. Когда он проходит, эволюция продолжается, но уровень новообразований постепенно идет на убыль. Если, например, речь идет о подклассе, то сначала перестают появляться новые порядки, потом новые семейства, потом новые роды, потом новые виды. Старые же постепенно вымирают.
Есть основания предполагать, что темпы эволюционного процесса всё время возрастают. Мы уже говорили об этом. Чтобы зеленые водоросли просто cозрели для выхода на сушу, потребовался миллиард лет. Чтобы сформировалось гигантское, невообразимое ранее, разнообразие покрытосеменных, оказалось достаточно 200 млн. лет. Это означает, что прогрессивно эволюционируют не просто сами растения. Прогрессивно эволюционирует их способность эволюционировать.
 На рисунке по вертикальной оси показана скорость видообразования (сколько новых видов образуется за единицу времени), а по горизонтальной – средняя продолжительность жизни одного отдельно взятого вида (млн. лет). Черными кружками показаны первые наземные растения, черными треугольниками – высшие споровые (папоротники, хвощи и плауны), черными квадратами – голосеменные, пустыми кружками - покрытосеменные. Как видите, в ходе эволюции скорость видообразования все время растет, продолжительность жизни отдельных видов сокращается. Современные голосеменные (обозначены красным) занимают в той системе промежуточное положение между споровыми и покрытосеменными, но располагаются намного ближе к первым, чем к последним. Источник рисунка - Niklas, K., B. Tiffney, et al. 1983. Patterns in vascular land plant diversification. Nature 303: 614–616.
На рисунке по вертикальной оси показана скорость видообразования (сколько новых видов образуется за единицу времени), а по горизонтальной – средняя продолжительность жизни одного отдельно взятого вида (млн. лет). Черными кружками показаны первые наземные растения, черными треугольниками – высшие споровые (папоротники, хвощи и плауны), черными квадратами – голосеменные, пустыми кружками - покрытосеменные. Как видите, в ходе эволюции скорость видообразования все время растет, продолжительность жизни отдельных видов сокращается. Современные голосеменные (обозначены красным) занимают в той системе промежуточное положение между споровыми и покрытосеменными, но располагаются намного ближе к первым, чем к последним. Источник рисунка - Niklas, K., B. Tiffney, et al. 1983. Patterns in vascular land plant diversification. Nature 303: 614–616.
Геном голосеменных: особенности и их значение для эволюции
Есть основания полагать, что изменение темпов эволюции связано с изменениями в устройстве и работе генома. Как они происходят? Современная наука постепенно начинает приоткрывать вековую завесу над этой тайной. В частности, некоторую пищу для размышлений дает сравнение геномов у голосеменных и цветковых. Скорость эволюции – это в значительной мере скорость появления мутаций. Если речь идет об эволюции генома, то ее можно измерить скоростью появления нуклеотидных замен в цепи ДНК. Анализ окаменелостей позволяет установить сроки расхождения отдельных филогенетических линий. Анализ последовательностей ДНК позволяет установить, сколько нуклеотидных замен произошло за это время в конкретных генах. Всё это вместе позволяет оценить скорость молекулярной эволюции. Сравнительный анализ голосеменных и цветковых проведен вот в этой очень серьезной работе: Amanda R. De La Torre, Zhen Li, Yves Van de Peer, and Par K. Ingvarsson. 2017. Contrasting Rates of Molecular Evolution and Patterns of Selection among Gymnosperms and Flowering Plants. Molecular Biology and Evolution. Vol. 34 (6): 1363–1377. У голосеменных скорость молекулярной эволюции (мутаций – замен нуклеотидов) в 7 раз ниже (в среднем), чем у цветковых. В 7 раз!!! Это приближает нас к пониманию того, почему саговники, гинкго, араукарии и даже сосны были примерно такими же, как сейчас, задолго до появления цветковых. Главных причин предположительно две. Первая лежит на поверхности. Это возраст начала половой репродукции. У голосеменных он в среднем многократно выше, чем у цветковых. Поэтому за единицу времени, например, за тысячу лет сменяется меньше поколений.
Вторая причина невооруженным глазом не видна, но она, по-видимому, важнее, и в значительной мере определяет первую. Чтобы произошло деление клетки, вся ее ядерная ДНК должна дублироваться. Чем ее больше, тем больше нужно для этого времени, вещества и энергии при прочих равных условиях. Так вот: у наших хвойных деревьев геном в 10, а то и в 100 раз больше, чем у наших лиственных.
 Если сравнивать голосеменные и цветковые в целом, то различие будет примерно в 3 раза. В отношении размера генома голосеменные (Gymnosperms) ближе к папоротникам (Ferns), но превосходят даже их. Разнообразие внутри каждой группы – огромное, причем, у цветковых (Angyosperms) оно даже больше, чем у голосеменных. Однако немногочисленные исключения лишь подтверждают общее правило. Когда-то давно предполагалось, что у голосеменных и генов в разы больше. Эта гипотеза не подтвердилась: число работающих (кодирующих белки) генов у всех семенных растений примерно одинаковое: 25-30 тыс. Всё остальное в геноме голосеменных – это по большей части генетический мусор, который они вынуждены копировать и перекопировать при каждом клеточном делении. Тут уже не до эволюции: дай бог выжить! Ссылка на источник рисунка.
Если сравнивать голосеменные и цветковые в целом, то различие будет примерно в 3 раза. В отношении размера генома голосеменные (Gymnosperms) ближе к папоротникам (Ferns), но превосходят даже их. Разнообразие внутри каждой группы – огромное, причем, у цветковых (Angyosperms) оно даже больше, чем у голосеменных. Однако немногочисленные исключения лишь подтверждают общее правило. Когда-то давно предполагалось, что у голосеменных и генов в разы больше. Эта гипотеза не подтвердилась: число работающих (кодирующих белки) генов у всех семенных растений примерно одинаковое: 25-30 тыс. Всё остальное в геноме голосеменных – это по большей части генетический мусор, который они вынуждены копировать и перекопировать при каждом клеточном делении. Тут уже не до эволюции: дай бог выжить! Ссылка на источник рисунка.
Раз уж мы заговорили о геномах и их структуре, остановимся на этом чуть подробнее. Геном водорослей – 5-7 млн. пар нуклеотидов. Это очень мало. Клетки с таким геномом делятся быстро. Организмы, из которых они состоят, отлично живут в привычной среде, но на прогресс и усложнение не замахиваются. Выход растений на сушу, который мы обсуждали выше, сопровождался многократным (примерно в 100 раз) увеличением размера генома, примерно до миллиарда пар нуклеотидов. Это, видимо, происходило по принципу цепной реакции: увеличение генома способствовало решению сложных задач, структурно-функциональное усложнение организмов требовало дальнейшего увеличения генома, это увеличение способствовало решению новых задач и т.д. Кстати, ровно то же самое происходило и у животных при переходе от беспозвоночных к позвоночным. Даже «цифра» получилась похожая: тот же самый миллиард. Увеличение размеров генома в обеих линиях (и у растений, и у животных), с одной стороны, сопровождалось усложнением его структуры. С другой стороны, происходило методом проб и ошибок.
Важнейшей областью таких проб и ошибок стал сам размер генома. Поразительно, но факт: в переходной от водорослей к семенным растениям группе подклассов (плауны + хвощи + папоротники) размер генома у разных видов варьирует в широченном диапазоне: от 100 млн. до 300 млрд. пар нуклеотидов! То же самое мы видим в переходных группах у животных: среди рыб и амфибий. Двигаться дальше по пути прогресса можно было только «разобравшись» с этим вопросом. Животные и растения «разобрались»: нашли золотую середину. Абсолютное большинство цветковых имеют размер генома в диапазоне от 0,5 до 5 млрд. (в среднем 2,7 млрд.). У высших животных диапазоны еще меньше: 1,3-2,5 млрд. у рептилий, 1,7-2,3 млрд. у птиц (в среднем 2,2 млрд.), 1,4-5,7 млрд. у млекопитающих. Человек, между прочим, располагается ровно посередине последнего диапазона – 3 млрд. пар.
Как выглядят в этой системе голосеменные? Размер их генома варьирует от 2 до 37 млрд. пар, в среднем 16 млрд. Это очень много. Гигантский размер генома голосеменных долго не давался ученым. Подступиться к нему удалось совсем недавно. Только в 2003 г. была завершена работа по секвенированию первого полного генома голосеменных на примере ели европейской, Picea abies. Дело было в Швеции. Результаты опубликованы: Nystedt B. et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497: 579-584. О масштабе этой работы говорит хотя бы число авторов. «Nystedt B. et al.» означает Nystedt B. и другие. Так вот, других тут не много и не мало, а 55 человек. Эта работа открыла новый этап в эволюционной и общей геномике растений. Авторы диагностировали у голосеменных новое явление, которое они образно назвали «genomic obesity = геномное ожирение».
Что это значит? Чтобы разобраться, начнем издалека. Во многих случаях важен не столько размер генома, сколько его структура. В эволюции многих растительных групп большое значение имеет полиплоидия, т.е. удвоение, утроение и т.д. целых геномов. При этом количество ДНК в клетке сразу же увеличивается вдвое, втрое и т.д. Это, в основном, сугубо позитивное для эволюции явление. У полиплоидов по сравнению с обычными диплоидами активней синтезируются белки, повышается общая пластичность и адаптивность, появляются возможности для диверсификации функций (из двух одинаковых генов один со временем начинает делать что-то другое и полезное). Среди современных цветковых 50-80% видов (по разным оценкам) – явные полиплоиды. А ведь есть еще много скрытых, у которых полиплоидизация произошла давно, поэтому идентифицируется не так надежно. Лишь у голосеменных полиплоиды – редкость: не более 5% видов, а у хвойных и того меньше – 1,5%. Значит, гигантские геномы у голосеменных появились явно не по этой причине.

Упрощенная филогения наземных растений с фокусом на WGD (whole-genome duplication = дупликацию полных геномов). Таких дупликаций (желтые ромбики) было немало, но все они были СОБЫТИЯМИ. Их так и называют - event, обозначая буквами древнегреческого алфавита: alpha-event, beta-event и т.д. Обратите внимание, что дупликация целого генома, как правило, предшествует важным эволюционным «прорывам»: оно было в начале диверсификации семенных (seed plants) (зета-событие), цветковых (flowering plants) (эпсилон-событие), однодомных (monocots) (тау-событие), двудомных (eudicots) (гамма-событие). В эволюции злаков (Zea, Oryza, Triticum) было две дупликации, сигма- и ро-события: одно до, другое после Cretaceous–Tertiary extinction event = массового вымирания на границе мезозоя и кайнозоя, всем известного как время гибели динозавров. Дупликации целого генома - важное отличие высших растений от высших животных: у последних ничего такого не известно. Полиполоидизация играла и играет огромную роль в эволюции цветковых. Скорее всего, это один из главных факторов их безоговорочной победы в борьбе за существование. Про голосеменные (gymnosperms) мы этого сказать не можем. На схеме обозначены два случая древней WGD. Оба относятся к хвойным и, как положено, локализованы в основании филогенетических ветвей: (1) семейства сосновые и (2) остальных хвойных. Однако они не вполне доказаны, некоторые исследователи сомневаются в их существовании. Поэтому греческими буквами они не обозначены. Случаи недавней полиплоидизцации также показаны на схеме. Однако этими несколькими случаями, собственно говоря, почти исчерпывается их перечень у голосеменных. Источник рисунка - Alix K, Gérard PR, Schwarzacher T, Heslop-Harrison JS (2017) Polyploidy and interspecific hybridization: partners for adaptation, speciation and evolution in plants. Ann Bot 120(2):183–194
Гигантский геном голосеменных сформировался совсем по-другому. Он на 80 и более процентов состоит из так называемых мобильных генетических элементов. Так называются небольшие фрагменты ДНК, которые могут самопроизвольно вырезаться из молекулы, копироваться, вставляться на новое место: методом copy-and-past. Они и составляют большую часть того генетического мусора, который я упоминал выше. На самом деле, это, конечно, некоторое преувеличение. Польза от них тоже есть, в том числе и для эволюции. Но по большей части это всё-таки балласт. Они накапливались в геноме голосеменных медленно и постепенно. Но за десятки миллионов лет накопилось их очень и очень много. Это и есть то самый геномный «жир», который обнаружили шведские ученые в геноме ели. Именно они делают геном голосеменных тяжелым, неповоротливым, нединамичным. Голосеменные, увы, плохо умеют от него избавляться. Возможно, в этом главная причина их поражения в борьбе за существование с цветковыми.
В геноме последних, конечно, тоже есть мобильные генетические элементы, но в разумном, полезном для «здоровья» количестве. Цветковые, похоже, изначально имели способность использовать излишки мобильных генетических элементов именно как балласт, т.е. «сжигать» излишний «жир» в своем геноме, «худеть» при необходимости и просто для профилактики. Эта способность относится не только к элиминации лишних мобильных элементов, но вообще к оптимизации размера. Ведь если полиплоидизация – важнейший фактор эволюции цветковых, а другой ее важнейший фактор – компактность генома, то главный принцип эволюции цветковых очевиден: сначала увеличить размер генома, потом «поработать» с ним, сформировать из излишков что-то новенькое и полезное, а от всего ненужного избавиться. Эффективно функционирующий механизм оптимизации размеров генома (“genome downsizing”) – это, возможно, главная причина грандиозного успеха цветковых.
Всё это были условно плохие новости про геном голосеменных (исключая гнетовые). Есть и хорошая. Она содержится вот в этой статье: Michael D. Crisp and Lyn G. Cook. 2011. Cenozoic extinctions account for the low diversity of extant gymnosperms compared with angiosperms. The New Phytologist, Vol. 192, No. 4, pp. 997-1009. Авторы проанализировали два филогенетических древа: голосеменных и цветковых. Оказалось, что средний возраст кайнозойских ответвлений у голосеменных был значительно меньше (32 млн. лет), чем у цветковых (50 млн. лет). Это означает, что в первую половину кайнозоя цветковые активно эволюционировали, в частности, появлялось много новых родов. Напротив, голосеменные в это время преимущественно вымирали. Они «подняли голову» лишь во второй половине кайнозоя, когда у них появились новые роды, а большинство современных видов и вовсе имеют возраст 5-10-15 млн. лет, причем, в некоторых родах видообразование было очень активным.
 С чем всё это связано? С изменениями климата в кайнозое. Современные научные представления о них показаны на рисунке из вот этой публикации: Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686–693. 50 млн. лет назад (начало эоцена) - это самый теплый период кайнозоя, когда почти по всей планете были распространены так называемые бореотропические леса. Они состояли почти исключительно из цветковых, которые процветали и доминировали, активно развивались. Голосеменные в это время были оттеснены цветковыми в приполярные и высокогорные рефугиумы (от лат. refugium — убежище), где они, впрочем, не просто выживали, а вырабатывали адаптации к сухому и холодному климату. Такой климат не заставил себя долго ждать. Он наступил в начале олигоцена, примерно 35 млн. лет назад. Голосеменные были к этому готовы. Тропические леса отступили к экватору. На их месте в умеренных широтах появились неморальные и бореальные леса, в которых происходил новый этап эволюции голосеменных. С середины миоцена (15 млн. лет назад) началось еще более серьезное похолодание, которое вызвало дальнейший рост разнообразия голосеменных, особенно на видовом уровне. Этот период продолжается до сих пор. Считается, что именно в это время у голосеменных возникли многие принципиальные инновации: листопадность, ксерофитная структура листа, разнообразные адаптации к распространению семян животными. Современные группы голосеменных – не дряхлеющие долгожители, они занимают разнообразные экологические ниши и вполне адаптивны. Во многих родах много настоящих и при этом очень близких, совсем молодых видов. Получается, не всё так грустно: у голосеменных еще есть «порох в пороховницах».
С чем всё это связано? С изменениями климата в кайнозое. Современные научные представления о них показаны на рисунке из вот этой публикации: Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686–693. 50 млн. лет назад (начало эоцена) - это самый теплый период кайнозоя, когда почти по всей планете были распространены так называемые бореотропические леса. Они состояли почти исключительно из цветковых, которые процветали и доминировали, активно развивались. Голосеменные в это время были оттеснены цветковыми в приполярные и высокогорные рефугиумы (от лат. refugium — убежище), где они, впрочем, не просто выживали, а вырабатывали адаптации к сухому и холодному климату. Такой климат не заставил себя долго ждать. Он наступил в начале олигоцена, примерно 35 млн. лет назад. Голосеменные были к этому готовы. Тропические леса отступили к экватору. На их месте в умеренных широтах появились неморальные и бореальные леса, в которых происходил новый этап эволюции голосеменных. С середины миоцена (15 млн. лет назад) началось еще более серьезное похолодание, которое вызвало дальнейший рост разнообразия голосеменных, особенно на видовом уровне. Этот период продолжается до сих пор. Считается, что именно в это время у голосеменных возникли многие принципиальные инновации: листопадность, ксерофитная структура листа, разнообразные адаптации к распространению семян животными. Современные группы голосеменных – не дряхлеющие долгожители, они занимают разнообразные экологические ниши и вполне адаптивны. Во многих родах много настоящих и при этом очень близких, совсем молодых видов. Получается, не всё так грустно: у голосеменных еще есть «порох в пороховницах».
Тем более, что говорить о голосеменных в целом – это всё равно, что обсуждать среднюю температуру по больнице. Их ведь не зря разделили на несколько независимых подклассов. Они очень разные. Если мы сразу начнем рассматривать эту разность в мелких подробностях, есть опасность, что не все пользователи успешно «въедут» в тему. Поэтому предлагаю поступить так. Сначала мы посмотрим на предков современных голосеменных, потом коротенько, буквально в 2-3 словах и 2-3 картинках очень-очень поверхностно познакомимся с 4 современными подклассами, а уже потом перейдем к их детальному сравнению.
Голосеменные подклассы: поверхностное знакомство
Знатоки и хранители русского языка сильно скривились, прочитав это словосочетание: голосеменные подклассы. А вот и зря. Мы ведь договорились, что класса голосеменные в современной систематике нет. Значит, нет у него и подклассов. Однако некоторые подклассы класса Equisetopsida мы можем неформально объединить именно по признаку голосемянности. Их, за невозможностью лучшего, мы и будем называть голосеменными подклассами.
Общепризнано, что в основании древа всех современных семенных растений находятся так называемые семенные папоротники, которые процветали в последнюю треть палеозойской эры. Их суть понятна из названия: это разноспоровые папоротники с семенами как органами размножения и расселения. Мегаспора, прорастающая прямо в мегаспорангии, есть и у некоторых обычных папоротников. Главной новацией семенных папоротников был интегумент – покров вокруг индивидуального мегаспорангия, который делает из этого мегаспорангия семяпочку, а впоследствии и семя. Семена у первых семенных папоротников располагались прямо на зеленых листьях.
 Семенные папоротники были широко распространены и активно участвовали в формировании каменного угля. Поэтому окаменевших остатков от них сохранилось довольно много. Они хорошо изучены. По ним сделаны реконструкции. Одна из них перед вами. Ссылка на источник рисунка.
Семенные папоротники были широко распространены и активно участвовали в формировании каменного угля. Поэтому окаменевших остатков от них сохранилось довольно много. Они хорошо изучены. По ним сделаны реконструкции. Одна из них перед вами. Ссылка на источник рисунка.
Это самый древний, примитивный вариант: листья чисто папоротниковидные, специализированных спорофиллов нет, мегаспорангии одиночные. Однако семенные папоротники были широко распространены и довольно разнообразны: это несколько порядков, до десятка семейств, не один десяток родов, сотни видов. Похоже, что уже в начале их эволюции обособились несколько перспективных групп, которые дали начало 5 современным подклассам семенных растений, 4 из которых традиционно относят к голосеменным.

Подкласс Cycadidae (саговниковые)
Чаще всего выглядят вот так (похожими на пальму, Dioon spinulosum; слева) или вот так (похожими на одревесневшую репку, которая, правда, не видна потому, что скрыта в почве, Zamia pygmea; справа). Листья имеют большие, папоротниковидные. Лишь генеративные органы выдают в них некую близость к привычным для нас голосеменным. Живут почти исключительно в тропиках. Ссылка на источник фотографии слева, ссылка на источник фотографии справа.

{kind=link}
{kind=link}
{kind=link}
{kind=link}


Подкласс Ginkgoidae (гинкговые)
Говорить о них во множественном числе как-то непривычно: в современной флоре нашей планеты они представлены единственным видом. Издалека, да и вблизи, этот вид похож на обычное лиственное дерево, даже отчасти плодовое. Живет в субтропиках. Ссылки на источники фотографий: слева, справа вверху.


Подкласс Pinidae (хвойные)
Это понятно без объяснений. Хотя, на самом деле, хвойные довольно разнообразны. Приведем пару фотографий, которые, возможно, удивят неискушенного пользователя (Phyllocladus asplenifolius слева, Podocarpus macrophyilla справа вверху). Живут, главным образом, в умеренном поясе, но есть и в тропиках. Ссылки на источники фотографий: слева, справа вверху.
{kind=link}



Подкласс Gnetidae (гнетовые)
Удивительная группа, главная ботаническая загадка на все времена. Кто-то похож на лиственные (Gnetum leyboldii, слева), кто-то на хвощ с ягодами (Ephedra equisetina, в центре), кто-то вообще ни на что не похож (Welwitschia mirabilis, справа). Живут в очень разных, в том числе, совершенно неожиданных местах. Ссылка на источники фотографий: слева, в центре, справа.
{kind=link}
{kind=link}
{kind=link}
Голосеменные подклассы: происхождение и родственные связи
Ну вот, теперь можно переходить к подробностям. Начнем с происхождения и родственных связей. В последние 20 лет о них судят исключительно по молекулярно-генетическим данным (ДНК). Сначала отдельные построения, полученные разными методами, бывало, противоречили друг другу и классическим (сделанным по морфологическим признакам) схемам. Однако прогресс был очень быстрый: совершенствовались методы, накапливалась информация. Последние молекулярные филогении по-прежнему во многом противоречат классическим, но уже вполне удовлетворительно согласуются между собой, что свидетельствует об их объективности. Приведем одну из них: Lu Y, Ran J-H, Guo D-M, Yang Z-Y, Wang X-Q (2014) Phylogeny and Divergence Times of Gymnosperms Inferred from Single-Copy Nuclear Genes. PLoSONE 9(9): e107679. doi:10.1371/journal.pone.0107679. Она, как видите, сделана нашими бывшими (или будущими) братьями – китайцами. Да, они производят теперь большую часть не только промышленной, но и научной продукции. А в области филогенетических исследований «Made in China» - это знак качества.

Итак, одна из последних филогенетических реконструкций, сделанная по двум монокопийным ядерным генам. Это очень надежный, проверенный метод. Филогения, естественно, калибрована по окаменелостям. В ней отражено не только родство разных групп, но и время их дивергенции (расхождения). Что мы видим? Мы видим, что ветви-подклассы последовательно отчленялись от общего ствола. Самая древняя ветвь (350 млн. лет назад) – саговниковые (на схеме это два семейства, Cycadaceae и Zamiaceae). Вторая по возрасту ветвь (340 млн. лет назад) – гинкговые (Ginkgoaceae). До этого места всё идет более или менее привычно. Дальше начинаются удивительные вещи. Следующая ветвь (чуть моложе гинкговых) – это хвойные, но не в полном составе, а лишь семейство сосновые (Pinaceae). Следующая ветвь (320 млн. лет назад) – гнетовые (порядок Gnetales). Последняя ветвь – все остальные хвойные (6 семейств от Sciadopityaceae до Cupressaceae).
Это поразительно, но это факт, подтвержденный и многими другими исследованиями: сосновые в филогенетическом отношении дальше от остальных хвойных, чем те и другие от гнетовых. Если признать это истиной в последней инстанции, то по правилам естественной систематики возможны лишь два варианта действий: (1) включить гнетовые в состав подкласса Pinidae (хвойные), (2) разделить этот подкласс хотя бы на два подкласса – сосновые и прочие, оставив гнетовые самостоятельным подклассом. Современная систематика растений уже, можно сказать, капитулировала под натиском молекулярных биологов: она активно меняет привычные схемы. Но случай с гнетовыми – просто вопиющий, невиданный и неслыханный. Допустить, что вельвичия ближе к сосне, чем, например, туя, никто из систематиков пока не готов. Это, можно сказать, последний бастион морфологической систематики, который она защищает грудью. Только через мой труп: написано на ее знамени. Поэтому, как говорится, запасаемся попкорном. Будем следить: чья возьмет. Пока же отметим, что в научной литературе всё чаще встречается деление хвойных на две неформальные группы: хвойные I (семейство Pinaceae) и хвойные II (остальные 6 семейств).
 В учебной литературе широко распространен вот такой способ представления филогении, который показывает не только возраст таксономических групп, но и их относительное участие в растительности нашей планеты. Толщина филогенетической ветви характеризует уровень ее процветания или упадка. Как видите, с конца юрского периода и до нашего времени абсолютно доминируют цветковые (темно-зеленый цвет). Голосеменные доминировали в конце палеозоя и первой половины мезозоя. Среди них много полностью вымерших таксонов, как очень древних, так и относительно молодых. Когда-то их было много, потом они полностью исчезли. Хорошо видны два цикла вымирания: конец полеозоя и конец мезозоя. Они совпадают с катастрофами планетарного масштаба, по которым, собственно, и проведены границы между эрами. Примитивные современные подклассы голосеменных появились в середине мезозоя, хвойные чуть позже. Лишь гнетовые заметно запоздали. Все современные подклассы знавали лучшие времена, особенно в середине мезозойской эры. Кайнозойское вымирание почти не коснулось хвойных, остальные подклассы заметно подсократились, особенно гинкговые. Источник рисунка - Linkies A, Graeber K, Knight C, Leubner-Metzger G (2010) The evolution of seeds. New Phytol 186:817–831.
В учебной литературе широко распространен вот такой способ представления филогении, который показывает не только возраст таксономических групп, но и их относительное участие в растительности нашей планеты. Толщина филогенетической ветви характеризует уровень ее процветания или упадка. Как видите, с конца юрского периода и до нашего времени абсолютно доминируют цветковые (темно-зеленый цвет). Голосеменные доминировали в конце палеозоя и первой половины мезозоя. Среди них много полностью вымерших таксонов, как очень древних, так и относительно молодых. Когда-то их было много, потом они полностью исчезли. Хорошо видны два цикла вымирания: конец полеозоя и конец мезозоя. Они совпадают с катастрофами планетарного масштаба, по которым, собственно, и проведены границы между эрами. Примитивные современные подклассы голосеменных появились в середине мезозоя, хвойные чуть позже. Лишь гнетовые заметно запоздали. Все современные подклассы знавали лучшие времена, особенно в середине мезозойской эры. Кайнозойское вымирание почти не коснулось хвойных, остальные подклассы заметно подсократились, особенно гинкговые. Источник рисунка - Linkies A, Graeber K, Knight C, Leubner-Metzger G (2010) The evolution of seeds. New Phytol 186:817–831.
 Посмотрим на эти же процессы немножко по-другому: не со стороны участия в экосистемах, а со стороны разнообразия, в данном случае, числа видов (вертикальная ось). На картинке видны некоторые интересные подробности. Так, во второй половине палеозоя доминировали плауны (Lycopsids), хвощи (Sphenopsids) и семенные папоротники (Pteridosperms). В мезозое доминирующее положение заняли папоротники (Ferns) и разные группы голосеменных. В конце мезозоя некоторые группы голосеменных полностью исчезли, доля других групп, а также папоротников, резко сократилась. Изменения происходили поразительно быстро. Цветковые заняли доминирующее положение буквально за 10-15 млн. лет в середине мелового периода. Обратите внимание, что разнообразие гинкговых никогда не было большим, а гнетовые тут вовсе не показаны (их всегда было мало). Источник рисунка - Knoll, A. 1986. Patterns of change in plant communities through geological time. Editors: J.A. Diamond and T.J. Case. In: Community Ecology. Harper & Row, New York. Chapter 7, pp. 126–141.
Посмотрим на эти же процессы немножко по-другому: не со стороны участия в экосистемах, а со стороны разнообразия, в данном случае, числа видов (вертикальная ось). На картинке видны некоторые интересные подробности. Так, во второй половине палеозоя доминировали плауны (Lycopsids), хвощи (Sphenopsids) и семенные папоротники (Pteridosperms). В мезозое доминирующее положение заняли папоротники (Ferns) и разные группы голосеменных. В конце мезозоя некоторые группы голосеменных полностью исчезли, доля других групп, а также папоротников, резко сократилась. Изменения происходили поразительно быстро. Цветковые заняли доминирующее положение буквально за 10-15 млн. лет в середине мелового периода. Обратите внимание, что разнообразие гинкговых никогда не было большим, а гнетовые тут вовсе не показаны (их всегда было мало). Источник рисунка - Knoll, A. 1986. Patterns of change in plant communities through geological time. Editors: J.A. Diamond and T.J. Case. In: Community Ecology. Harper & Row, New York. Chapter 7, pp. 126–141.
Голосеменные подклассы: размер и структура генома
Хромосомы у голосеменных крупные, хорошо видны под микроскопом. Поэтому изучены основательно. Их число довольно изменчиво: 2 n от 14 до 66. Источника изменчивости условно два: обычное разнообразие и полиплоидия. Начнем с последнего. Явная, бесспорная, «молодая» полиплоидия – большая редкость у голосеменных (4-5% видов). Она есть только у хвойных (всего несколько видов, в том числе, гексаплоид Sequoia, 6 n = 66), остальные относятся к роду Ephedra из гнетовых (более половины из примерно 50 видов).
Обычная изменчивость (если исключить явные полиплоиды) – заметно меньше: 2 n от 14 до 42. С принадлежностью к подклассам это почти не связано. Больше связано с принадлежностью к семействам. Например, в семействе Cupressaceae (кипарисовые) число хромосом у всех диплоидных родов и видов одинаковое (2 n = 22), а в семействе Podocarpaceae (подокарповые) из того же подкласса хвойные – сильно варьирует (2 n от 18 до 38). В семействах с большой изменчивостью она есть и внутри родов: например, у Podocarpus 2 n от 20 до 38.
Выше, когда мы обсуждали размер генома, то измеряли его в парах нуклеотидов. Такая информация есть пока далеко не для всех видов. Другие методы позволяют оценить размер генома в граммах, а точнее в пикограммах (пг = одна триллионная грамма). Удачно получилось, что 1 пг = 978 млн. пар нуклеотидов, т.е. примерно 1 млрд. Впрочем, эти величины всё равно невозможно себе представить. Поэтому не будем и пытаться: нам ведь для наших целей важны не столько абсолютные, сколько относительные показатели. Огромный размер генома голосеменных, равно как и его значение, мы уже обсуждали. Абсолютный диапазон разнообразия от 4,5 пг у одного из видов Gnetum до 72 пг у одного из видов Pinus.
 Есть ли тут различия между подклассами? Есть. Они показаны на рисунке. Вертикальная ось – это 79 родов голосеменных, горизонтальная ось – средний размер одного диплоидного генома (2 n, за исключением нескольких полиплоидов) у каждого рода. Внутри подклассов различия большие: 2-кратные у саговниковых, 4-кратные у гнетовых, 8-кратные у хвойных. По среднему размеру генома (45 пг) сильно выделяются саговниковые (желтые линии). Хвойные (голубые линии) заметно уступают им (27 пг). Гинкго (зеленая линия) находится на среднем хвойном уровне (23,5 пг). Самый маленький геном (17 пг) у гнетовых (красные линии). И это при том, что к ним относится род эфедра (средний размер генома 31,4 пг), где много полиплоидов (есть даже октаплоиды, 8 n). Как видите, размер генома убывает от древних подклассов к молодым. С сокращением размера генома увеличивается скорость эволюции и вообще эволюционный потенциал. Источник информации для рисунка - X.-Q. Wang, J.-H. Ran. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution. 75 (2014) 24–40.
Есть ли тут различия между подклассами? Есть. Они показаны на рисунке. Вертикальная ось – это 79 родов голосеменных, горизонтальная ось – средний размер одного диплоидного генома (2 n, за исключением нескольких полиплоидов) у каждого рода. Внутри подклассов различия большие: 2-кратные у саговниковых, 4-кратные у гнетовых, 8-кратные у хвойных. По среднему размеру генома (45 пг) сильно выделяются саговниковые (желтые линии). Хвойные (голубые линии) заметно уступают им (27 пг). Гинкго (зеленая линия) находится на среднем хвойном уровне (23,5 пг). Самый маленький геном (17 пг) у гнетовых (красные линии). И это при том, что к ним относится род эфедра (средний размер генома 31,4 пг), где много полиплоидов (есть даже октаплоиды, 8 n). Как видите, размер генома убывает от древних подклассов к молодым. С сокращением размера генома увеличивается скорость эволюции и вообще эволюционный потенциал. Источник информации для рисунка - X.-Q. Wang, J.-H. Ran. Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution. 75 (2014) 24–40.
Геном простых существ, например, бактерий, устроен просто: большая его часть, это последовательности ДНК, кодирующие белок: почти ничего «лишнего». Геном семенных растений, впрочем, как и геном позвоночных животных, многократно сложнее. Например, у человека до 75% генома – это вообще не гены, а так называемые межгенные промежутки. Они включают, в основном, разные повторы, копии и фрагменты непонятного назначения. Среди оставшихся 25% белок-кодирующие последовательности (они называются экзонами) – это от силы 1-2%. Всё остальное – интроны. Они ничего не кодируют. В «сухом остатке» количество белок-кодирующей ДНК в клетке человека всего лишь в 3 раза больше, чем у среднестатистической бактерии. Вот вам и венец творения! Это, конечно, не означает, что 99% человеческого генома есть генетический мусор. Науке уже многое известно о важных, главным образом, регуляторных, функциях некодирующей ДНК. Но и конкретного мусора там тоже немало.
 Вернемся к семенным растениям. Как разные их группы выглядят, например, по размеру некодирующей части генов (интронов). Здесь, в основном, действует простая закономерность: чем меньше геном, тем меньше интроны. Оно и понятно. Ведь главный источник увеличения интронов – всё те же мобильные генетические элементы: они, похоже, одинаково активно «плодятся» и в межгенной ДНК, и в некодирующей части генов. Посмотрим на картинку. Оранжевым цветом на ней показаны цветковые (рис, кукуруза и др.), фиолетовым – голосеменные: гинкго (G. biloba), сосна (P. taeda) и гнетум (G. montanum). Серыми кружками показан размер генома, линейными диаграммами – длина интронов. Как видите, оба показателя существенно снижаются от гинкговых к хвойным, затем к гнетовым, затем к примитивным цветковым (Amborella trichopoda), наконец, стабилизируются у большинства цветковых. Очевидно, что в ходе прогрессивной эволюции растения учатся бороться не только с общим, но и конкретно с внутренним (интроны) геномным «ожирением». Источник рисунка - Wan T, Liu Z M, Li L F, et al. A genome for gnetophytes and early evolution of seed plants. Nature plants, 2018, 4(2): 82-89.
Вернемся к семенным растениям. Как разные их группы выглядят, например, по размеру некодирующей части генов (интронов). Здесь, в основном, действует простая закономерность: чем меньше геном, тем меньше интроны. Оно и понятно. Ведь главный источник увеличения интронов – всё те же мобильные генетические элементы: они, похоже, одинаково активно «плодятся» и в межгенной ДНК, и в некодирующей части генов. Посмотрим на картинку. Оранжевым цветом на ней показаны цветковые (рис, кукуруза и др.), фиолетовым – голосеменные: гинкго (G. biloba), сосна (P. taeda) и гнетум (G. montanum). Серыми кружками показан размер генома, линейными диаграммами – длина интронов. Как видите, оба показателя существенно снижаются от гинкговых к хвойным, затем к гнетовым, затем к примитивным цветковым (Amborella trichopoda), наконец, стабилизируются у большинства цветковых. Очевидно, что в ходе прогрессивной эволюции растения учатся бороться не только с общим, но и конкретно с внутренним (интроны) геномным «ожирением». Источник рисунка - Wan T, Liu Z M, Li L F, et al. A genome for gnetophytes and early evolution of seed plants. Nature plants, 2018, 4(2): 82-89.

Геномная эволюция семенных растений изучена пока не очень глубоко и основательно. То, что известно, суммировано вот на этом рисунке. Показано филогенетическое древо 7 основных групп семенных растений. Снизу вверх это саговниковые, гинкговые, хвойные, гнетовые, примитивные цветковые, однодольные и двудольные. Звездами показано удвоение целого генома: красная звезда – доказанное (в начале эволюции цветковых), серыми – гипотетические (в начале эволюции семенных растений, в начале эволюции голосеменных и в начале эволюции примитивных голосеменных: саговниковые + гинкговые). Стрелками разного цвета показаны главные события эволюции: генома. Синий цвет (семенные растения в целом): появление самой «идеи» повторов, формирование их основных типов; появление и эволюция интронов. Оранжевый цвет (голосеменные в целом): большой размер генома за счет активной экспансии мобильных генетических элементов и низкого уровня их элиминации (генное «ожирение»); медленная эволюция. Зеленый цвет (цветковые): оптимальный размер генома, его высокая динамичность за счет активного размножения мобильных генетических элементов и такой же активной их элиминации; высокая скорость эволюции. Черный цвет (подкласс гнетовые): сокращение общего размера генома и конкретно длины интронов, быстрая эволюция в оригинальном направлении. Фиолетовый цвет (общие черты гнетовых и примитивных цветковых): оптимизация размера генома, его высокая динамичность, похожая структура интронов. Источник рисунка - Wan T, Liu Z M, Li L F, et al. A genome for gnetophytes and early evolution of seed plants. Nature plants, 2018, 4(2): 82-89.
Голосеменные подклассы: современное разнообразие и распространение
Перейдем к современному разнообразию. Оно представлено в таблице по данным самого авторитетного источника: Christenhusz, M.J.M. et al. (2011) A new classification and linear sequence of extant gymnosperms. Phytotaxa 19: 55–70.

Как видите, голосеменные подклассы очень разные. Уникальны гинкговые, представленные единственным видом. Перманентно процветают сосновые: уже примерно в течение 250 млн. лет. Удивляют саговниковые: несмотря на рекордную древность, обильны видами. Как и во всем, ни на что не похожи гнетовые: по одному роду в порядке, зато видов довольно много. Еще лучше эти характерные закономерности видны в таблице другого типа.

Более или менее гармонично структура разнообразия выглядит лишь у хвойных. Но это зависит от того, с чем сравнивать. Если с другими голосеменными, то да. Ну а если с цветковыми, то это, конечно, «слезы». Очень интересна ситуация с гнетовыми и саговниковыми. Число видов на род у них в 2-3 раза больше, чем у хвойных. Когда высших таксонов мало, а видов много – это явный признак возрождения после незавершенного вымирания. Трудно сказать, с чем это связано. Особенно если учесть, что живут они в самых разнообразных условиях по всему земному шару. Но и закономерностью это не назовешь: гинкговые, например, возрождаться не расположены. И это при том, что их единственный вид на удивление устойчив к современным болезням и вредителям. Видимо, потому, что его патогены и его вредители давно вымерли. Тут можно сказать только одно: неисповедимы пути эволюции.
Как это всё распределено по нашей планете? Из 83 родов более половины (45) есть в Азии, 31 в Австралии. Подавляющее большинство монотипых родов (значит, древних, реликтовых) найдены на этих двух континентах. Следовательно, азиатско-австралийский регион есть центр разнообразия и, скорее всего, центр происхождения голосеменных. На остальных континентах разнообразия меньше, хотя достоверно известно немало родов, которые возникли, например, в Африке или Америке. Всё это интересные и важные вопросы, но здесь мы не будем их рассматривать. Рассмотрим потом, когда будем разбираться с конкретными подклассами, порядками, семействами, родами и видами.
По занимаемому ареалу голосеменные уступают цветковым. Но не так уж сильно, как можно было предположить, исходя из их реликтового статуса. Они распространены от южных окраин южных до северных окраин северных материков, от дождевых лесов на экваторе до верхней границы древесной растительности в горах. Карту давать не будем, но скажем, что ареал голосеменных – это почти вся наша планета. Климатические предпочтения голосеменных чрезвычайно разнообразны.
 Приведем диаграмму, построенную по данным вот этой совсем свежей работы: Walas, L., et al. Sexual systems in gymnosperms: A review. Basic and Applied Ecology (2017), V.31: 1-9. Как видите, голосеменные есть во всех типах климата, кроме полярного. Мы привыкли думать, что их дом – это, главным образом, нетропические области. Здесь они, действительно, на виду и доминируют. Однако число видов в тропиках не меньше. Просто они не так заметны среди «буйства» покрытосеменных. Это, в первую очередь, все саговниковые и один из родов гнетовых, а также некоторые хвойные. В бореальной зоне голосеменных совсем мало: немногим больше сотни. Это почти исключительно хвойные. Если сопоставить число видов в обычном и сухом климате, то в последнем их будет в разы меньше. Это не потому, что голосеменные как-то по-особенному не любят сухость. Скорее наоборот. Просто в сухих регионах флора вообще многократно беднее. Из голосеменных там распространены, в основном, хвойные, а также два рода гнетовых.
Приведем диаграмму, построенную по данным вот этой совсем свежей работы: Walas, L., et al. Sexual systems in gymnosperms: A review. Basic and Applied Ecology (2017), V.31: 1-9. Как видите, голосеменные есть во всех типах климата, кроме полярного. Мы привыкли думать, что их дом – это, главным образом, нетропические области. Здесь они, действительно, на виду и доминируют. Однако число видов в тропиках не меньше. Просто они не так заметны среди «буйства» покрытосеменных. Это, в первую очередь, все саговниковые и один из родов гнетовых, а также некоторые хвойные. В бореальной зоне голосеменных совсем мало: немногим больше сотни. Это почти исключительно хвойные. Если сопоставить число видов в обычном и сухом климате, то в последнем их будет в разы меньше. Это не потому, что голосеменные как-то по-особенному не любят сухость. Скорее наоборот. Просто в сухих регионах флора вообще многократно беднее. Из голосеменных там распространены, в основном, хвойные, а также два рода гнетовых.
Это была флора голосеменных. Теперь несколько слов о роли голосеменных в составе растительности. Их доля максимальная в бореальной зоне северного полушария, по направлению к экватору она быстро снижается, по направлению от экватора на юг снова повышается, но не так сильно, как в северном полушарии. То же самое и в горах: максимум в верхней части лесного пояса, минимум – в нижней. Как выглядят в этой системе отдельные подклассы? Коснемся вскользь лишь некоторых примеров, где голосеменные не просто участвуют, а играют ключевую роль в растительных сообществах. Поэтому гинкговые мы пропустим: их, можно сказать, вообще нет в дикой природе.
Саговниковые в современном мире нечасто образуют сплошные заросли. Однако изредка такое встречается.
 На снимке Cycas revoluta на южных японских островах Рюкю. Здесь, особенно на побережьях, этот вид часто выступает фоновым растением. Ссылка на источник фотографии.
На снимке Cycas revoluta на южных японских островах Рюкю. Здесь, особенно на побережьях, этот вид часто выступает фоновым растением. Ссылка на источник фотографии.

Еще один представитель саговниковых, Encephalartos lanatus, обычен в невысоких суховатых горах южной Африки. Ссылка на источник фотографи.
{kind=link}

Последний пример. Macrozamia communis из Австралии доминирует в подлеске сухих эвкалиптовых лесов. Ссылка на источник фотографии.
{kind=link}
Гнетовые никогда не занимали сколько-нибудь заметного места в земной растительности. Тем не менее, есть такие места, где они образуют основу ландшафта и создают его лицо.

Ephedra dahurica в сухой хакасской степи. Это южная Сибирь, довольно жесткие условия, резко континентальный климат. Однако эфедра, как видите, очень обильна, буквально цветет и пахнет. Фото Сергея Драгана. Ссылка на источник фотографии.

Еще более суровое место – совершенно безводная пустыня Намиб в Юго-Западной Африке. Однако для Welwitschia mirabilis это не помеха. Конкурентов у нее тут на обозримом расстоянии просто нет. Ссылка на источник фотографии.
{kind=link}
О хвойных. Что такое тайга – зеленое море хвойного леса в бореальной зоне северного полушария – вы отлично знаете. Показывать не будем. Однако подавляющее большинство видов хвойных живут отнюдь не в тайге, а в южной половине умеренного пояса и субтропических горах, где они также могут быть деревьями первой величины и доминировать. О них мы подробно поговорим, когда будем обсуждать хвойные.
 Здесь приведем лишь один пример – дождевые хвойные леса на тихоокеанском побережье Северной Америки: от юга Аляски до севера Калифорнии. Это чуть ли не единственный случай, когда природные условия очень благоприятные, но голосеменные, несмотря на это, доминируют над цветковыми. Самые продуктивные в мире леса образованы не одним десятком хвойных видов. Обычная (не рекордная) высота деревьев здесь – 60-70 м. К этому уникальному биому мы будем возвращаться еще не раз при описании отдельных видов. Ссылка на источник фотографии.
Здесь приведем лишь один пример – дождевые хвойные леса на тихоокеанском побережье Северной Америки: от юга Аляски до севера Калифорнии. Это чуть ли не единственный случай, когда природные условия очень благоприятные, но голосеменные, несмотря на это, доминируют над цветковыми. Самые продуктивные в мире леса образованы не одним десятком хвойных видов. Обычная (не рекордная) высота деревьев здесь – 60-70 м. К этому уникальному биому мы будем возвращаться еще не раз при описании отдельных видов. Ссылка на источник фотографии.
{kind=link}

В тропиках хвойных не меньше, чем в умеренном поясе. Они здесь, как правило, являются не доминантами, а ингридиентами, т.е. изредка встречаются среди лиственных. Тем не менее, обширные хвойные леса возможны и в тропиках. Опять же приведем лишь один пример: Araucaria angustifolia в Бразилии. Ссылка на источник фотографии.
{kind=link}
Голосеменные подклассы: вегетативное тело
О размере тела и формах роста. Травянистых форм среди современных голосеменных нет. В прошлом они были, но вымерли. Древесные растения обычно делят по высоте не в линейной, а в экспоненциальной шкале. (1) До 25 см, как бы кустарнички и низкие стланцы. В холодных областях их на зиму прикрывает снег, в тёплых на период засухи - отмершие остатки растений. (2) До 3 м, как бы кустарники и высокие стланцы. Они часто живут в подлеске, под защитой более высоких древесных растений, либо просто в приземном слое воздуха, где ветер не так силен. (3) Более 3 м, собственно деревья. У них почки и концевые побеги, предназначенные для переживания неблагоприятного периода, расположены высоко над землёй, в совершенно открытой атмосфере.
 Для демонстрации разнообразия еще раз воспользуемся информацией из уже упомянутой выше статьи: Walas, L., et al. Sexual systems in gymnosperms: A review. Basic and Applied Ecology (2017), V.31: 1-9. Как видите, среди голосеменных явно преобладают настоящие деревья. Это гинкго, большинство хвойных, мало саговниковых, еще меньше гнетовых. Наоборот, совсем низких растений мало. Но они есть во всех подклассах, естественно, кроме гинкговых. Это либо виды из подлеска, либо из суровых условий (болота, высокогорья). Фотографии здесь приводить не будем. Приведем их в соответствующих разделах.
Для демонстрации разнообразия еще раз воспользуемся информацией из уже упомянутой выше статьи: Walas, L., et al. Sexual systems in gymnosperms: A review. Basic and Applied Ecology (2017), V.31: 1-9. Как видите, среди голосеменных явно преобладают настоящие деревья. Это гинкго, большинство хвойных, мало саговниковых, еще меньше гнетовых. Наоборот, совсем низких растений мало. Но они есть во всех подклассах, естественно, кроме гинкговых. Это либо виды из подлеска, либо из суровых условий (болота, высокогорья). Фотографии здесь приводить не будем. Приведем их в соответствующих разделах.

Отдельно выделяются лианы, высота которых больше зависит не от них, а от наличия и высоты опоры. Среди голосеменных это редкость и представлены они почти на 100% родом Gnetum, который и дал название подклассу гнетовые. Выглядит это примерно вот так. На снимке – ствол Gnetum ula из Индии. Ссылка на источник фотографии.
 Еще один случай, когда размер растения не определяет его положение относительно земли, это эпифиты – растения, которые прикреплены к другим растениям, но не паразитируют на них. Удивительно, но факт: среди голосеменных есть и такие. Это два вида Zamia из саговниковых. Они живут в дождевых тропических лесах Центральной и Южной Америки. Эпифитов тут не много, а очень много. Вот наши замии и поддались этой всеобщей моде. На снимке Z. pseudoparasitica (с папоротниковидными листьями). Ссылка на источник фотографии.
Еще один случай, когда размер растения не определяет его положение относительно земли, это эпифиты – растения, которые прикреплены к другим растениям, но не паразитируют на них. Удивительно, но факт: среди голосеменных есть и такие. Это два вида Zamia из саговниковых. Они живут в дождевых тропических лесах Центральной и Южной Америки. Эпифитов тут не много, а очень много. Вот наши замии и поддались этой всеобщей моде. На снимке Z. pseudoparasitica (с папоротниковидными листьями). Ссылка на источник фотографии.
{kind=link}
Все голосеменные растения многолетние, притом, древесные. Поэтому для них очень важен ствол. Оказывается, устроен он принципиально по-разному в разных подклассах. Все мы привыкли видеть его таким, как у хвойных: полнодревесное бревно, из которого получаются отличные пиломатериалы.

Действительно, хвойные – это так называемая пикноксильная (от греч. pyknos - плотный и xylon - древесина) линия эволюции, в которой кора и сердцевина относительно тонкие, а вторичная древесина мощная и плотная. Сюда же относятся гинкговые и часть гнетовых. Латеральная меристема – камбий – ежегодно образует новый слой древесины, в результате чего большую часть ствола занимает единый массив этой ткани, состоящей почти исключительно из трахеид. Конструкция получается очень прочная и позволяет деревьям вырастать до большой высоты. Ее поперечный срез представлен на рисунке справа. Однако есть еще и маноксильная (от греч. manos - рыхлый, неплотный) линия эволюции: все саговниковые + вельвичия из гнетовых. У них стебли имеют толстую сердцевину и кору, но слабо развитую и при этом рыхлую вторичную древесину. Трахеид в ней не так уж много. Зато много паренхимы: живых тонкостенных округлых клеток, содержащих много воды. Кстати, это одна из причин, по которым саговниковые не живут за пределами тропиков: мороз физически разрушает их древесину. Поперечный срез мы видим на рисунке слева. Есть несколько способов образования такой древесины. Мы не будем вдаваться в эти подробности. Отметим лишь, что механической прочности ствола, да и эффективному проведению воды, всё это отнюдь не способствует. Поэтому для саговниковых даже высота 5-7 м – большая редкость. Ссылка на источник рисунка.
 Как уже говорилось выше, проводящими воду элементами в древесине голосеменных являются трахеиды – длинные веретеновидные клетки с относительно узкими просветами и без сквозной перфорации. Из этого правила есть исключение – гнетовые. В их древесине присутствуют настоящие сосуды, такие же, как у покрытосеменных. На снимке поперечный срез через древесину гнетума. Хорошо видны и сосуды, и трахеиды. Очень похоже на обычную березу. Ссылка на источние рисунка.
Как уже говорилось выше, проводящими воду элементами в древесине голосеменных являются трахеиды – длинные веретеновидные клетки с относительно узкими просветами и без сквозной перфорации. Из этого правила есть исключение – гнетовые. В их древесине присутствуют настоящие сосуды, такие же, как у покрытосеменных. На снимке поперечный срез через древесину гнетума. Хорошо видны и сосуды, и трахеиды. Очень похоже на обычную березу. Ссылка на источние рисунка.

Листья у голосеменных, пожалуй, не менее разнообразны, чем у цветковых. Их можно условно разделить на три линии: макро-, мезо- и микрофильные. Макрофильная линия – это, конечно, саговниковые. Сразу видно, что это ближайшие и прямые потомки семенных папоротников. На снимке южноафриканский Encephalartos longifolius. Ссылка на источник фотографии.
{kind=link}

Мезофильная линия – это гинкговые. Их листья не крупные, но и не хвойные. Если на что-то и похожи, то опять же на листья некоторых папоротников. Не зря ведь в Англии гинкго называют Maidenhair tree, что дословно переводится как «дерево девичьих волос». Дело в том, что Maidenhair – английское название травянистого папоротника адиантума, или венерина волоса (Adiantum capillus-veneris), который популярен и у нас в комнатной культуре. Листочки этого папоротника похожи на листья гинкго. Поэтому англичане, не долго думая, просто добавили к его названию слово «tree». Ссылка на источник фотографии.
{kind=link}
Листья гнетовых разные до крайности. Но мы все их уже видели. Показывать по второму разу не будем. Листья хвойных дали им называние. Все вы отлично их знаете. Показывать опять же не будем.
 Хотя хвойные тоже не так просты в этом смысле, как кажется. Вот это, например, Agathis robusta с Новой Гвинеи. Ну и где у него хвоя? Ссылка на источник фотографии.
Хотя хвойные тоже не так просты в этом смысле, как кажется. Вот это, например, Agathis robusta с Новой Гвинеи. Ну и где у него хвоя? Ссылка на источник фотографии.
{kind=link}
Голосеменные подклассы: разделение полов
Перейдем к половой репродукции и начнем с разделения полов. Растения принципиально отличаются от животных тем, что они по большей части обоеполы: каждая особь образует и мужские, и женские репродуктивные структуры. Понятно почему: растения неподвижны, они не могут активно искать полового партнера. Проблему желательности перекрестного оплодотворения они решают другим способом: формированием разнообразных механизмов, ограничивающих самооплодотворение. В этой области огромных успехов достигли покрытосеменные. Поэтому среди них абсолютно преобладают виды с обоеполыми цветками, на худой конец, однодомные раздельнополые (тычиночные и пестичные цветки образуются на одной особи). Двудомных видов (мужские цветки на одних особях, женские – на других) среди покрытосеменных не более 6%.
У голосеменных ситуация совсем другая: доля двудомных видов в 10 с лишним раз больше – 65% (здесь далее приведены результаты из уже упомянутой выше работы: Walas, L., et al. (2017) Sexual systems in gymnosperms: A review. Basic and Applied Ecology). Двудомные виды относятся к 42 родам (ровно 50%). Три из четырех подклассов (все, кроме хвойных) полностью двудомны. Среди хвойных двудомными являются около 40% видов. Очевидно, что причина такого широкого распространения двудомности среди голосеменных – это их примитивность, в частности, отсутствие эффективных генетических механизмов автостерильности. В таких условиях двудомность – главное средство избежать самооплодотворения. Способы определения пола изучены лишь у некоторых видов голосеменных. Кое-где найдены настоящие половые хромосомы, как у животных.
Голосеменные подклассы: стробилы и споры
Все не-цветковые растения объединены понятием «стробил». Так называют собрание спороносных листьев, микро- и мегаспорофиллов. Первые собраны в микро- или мужские, последние в мега- или женские стробилы. Современная наука склоняется к тому, что стробилы голосеменных сформировались уже после того, как произошло обособление 4 основных филогенетических линий, которые мы считаем относящимися к голосеменным. В каждой из этих линий стробилы возникли независимо просто потому, что это эффективная форма организации спорофиллов. Вполне естественно, что в разных подклассах стробилы устроены по-разному.
 Простой исходный вариант – ось со спорофиллами - сохранился лишь у саговниковых. На переднем плане микро- на заднем мегастробилы саговника поникающего (Cycas revoluta). Они, как видите, одиночные, довольно крупные, длиной до 1 м. При этом микростробилы микроразмерами отнюдь не отличаются. Наоборот, они часто (как в данном случае) крупнее мегастробилов. Ссылка на источник фотографии.
Простой исходный вариант – ось со спорофиллами - сохранился лишь у саговниковых. На переднем плане микро- на заднем мегастробилы саговника поникающего (Cycas revoluta). Они, как видите, одиночные, довольно крупные, длиной до 1 м. При этом микростробилы микроразмерами отнюдь не отличаются. Наоборот, они часто (как в данном случае) крупнее мегастробилов. Ссылка на источник фотографии.
{kind=link}

У остальных голосеменных более или менее похожими на исходный вариант остались лишь микростробилы. Варьирует их размер, причем, исключительно в сторону уменьшения, если брать саговниковые за точку отсчета. На снимке микростробилы туи западной. Их размер не более 2,5 мм. Ссылка на источник фотографии.
Микроспорангии производят пыльцу: начавший развитие мужской гаметофит в оболочке микроспоры. Только начавший: это принципиальное отличие от цветковых, у которых пыльца выходит на свободу с уже готовыми мужскими гаметами. У голосеменных такого не бывает: спермии всегда образуются уже потом, после вылета пыльцы из мегаспорангия. Пыльца не зря так называется. Она очень мелкая, пылевидная, ибо должна не просто разноситься ветром, но и парить в восходящих токах теплого воздуха при полном безветрии. При этом в ней должно содержаться всё необходимое. Поэтому размер пыльцы у всех высших растений примерно одинаковый: варьирует в диапазоне от 10 до 100 мкм, в том числе у голосеменных от 10 до 60 мкм. С «продвинутостью» подклассов размер пыльцы, можно сказать, не связан. Да и внутри подклассов он варьирует слабо.
Форма пыльцевых зерен, наоборот, различается очень сильно. Не только между крупными таксонами, но и между родами, иногда даже между видами. На этом основан так называемый спорово-пыльцевой анализ. Пыльца имеет очень устойчивые к разрушению временем покровы. Поэтому очень долго сохраняется в горных породах. По составу пыльцы в каком-то земном слое, можно довольно точно определить, какая растительность была во время формирования этого слоя.
 Основные типы пыльцевых зерен у голосеменных. A – саговниковые: пыльцевое зерно по форме похоже на футбольный мяч, из которого выкачали весь воздух. B – гинкго: узбекская лепешка. С–F – хвойные: как видите, их пыльца довольно разнообразная; есть похожая на пыльцу саговниковых, есть с очень характерной формой. G–J – гнетовые: их пыльца больше похожа на пыльцу цветковых растений. Источник рисунка - Fernando DD, Quinn CR, Brenner ED, Owens JN (2010) Male gametophyte development and evolution in extant gymnosperms. Int J Plant Dev Bio 4: 47–63
Основные типы пыльцевых зерен у голосеменных. A – саговниковые: пыльцевое зерно по форме похоже на футбольный мяч, из которого выкачали весь воздух. B – гинкго: узбекская лепешка. С–F – хвойные: как видите, их пыльца довольно разнообразная; есть похожая на пыльцу саговниковых, есть с очень характерной формой. G–J – гнетовые: их пыльца больше похожа на пыльцу цветковых растений. Источник рисунка - Fernando DD, Quinn CR, Brenner ED, Owens JN (2010) Male gametophyte development and evolution in extant gymnosperms. Int J Plant Dev Bio 4: 47–63
Пусть наша пыльца пока полетает, а мы вернемся у мегастробилам, чтобы обсудить их развитие до того, как они будут готовы принять пыльцу. Мегастробилы, в отличие от микростробилов, на удивление разнообразны. Самый древний, производный напрямую от семенных папоротников, примитивный вариант характерен для единственного рода саговник (Cycas) из подкласса саговниковые.

Мегастробил саговника мы уже видели издалека. Теперь рассмотрим его вблизи. Это примерно половина пути от зеленых мегаспорофиллов семенных папоротников к обычным чешуям мегастробила, какие есть у всех остальных голосеменных. Мегастробил очень рыхлый. Микроспорофиллы – не зеленые, но перистые, как бы дифференцированные на черешок и сильно рассеченную пластинку. На черешках в основании пластинок сидят семяпочки. Если приглядеться, то их можно заметить. Фото Юлии Борисовой. Ссылка на источник фотографии.
{kind=link}

Очевидно, что перистые листоподобные мегаспорофиллы – это явный анахронизм, излишество и помеха для опыления. Поэтому даже в самом роде Cycas хорошо прослеживается эволюция в сторону редукции перистых мегаспорофиллов. Современные виды можно расположить, например, вот в такой ряд: (A) C. revoluta, (B) C. rumphii, (C) C. circinalis. Источник рисунка - Veit Martin Dörken & Brigitte Rozynek (2013). Proliferated megasporangiate strobili of Zamia furfuracea (Zamiaceae, Cycadales) and its possible evolutionary implications for the origin of cycad-megasporophylls. Palaeodiversity 6: 135–147.
Редукция мегаспорофиллов – это лишь часть вопроса. При всей своей голосемянности голосеменные уже на заре эволюции «осознали», что для эффективного семенного размножения семяпочки и всё, что из них формируется, должно быть более или менее приоткрыто для внешней среды лишь на двух этапах развития: при опылении и после созревания семян. В течение всего остального времени их хорошо бы надежно «запечатать» от греха подальше. Так появилась ШИШКА, характерная для большинства голосеменных: компактная, покрытая более или менее жесткими чешуями, к которым прикреплены и под которыми спрятаны семяпочки, впоследствии – созревающие семена.
 Термин «шишка» в строгом смысле применим не ко всем голосеменным. Почему – будет сказано чуть ниже. Однако уже все роды саговниковых, кроме Cycas, имеют компактный мегастробил с жесткими чешуями. На снимке показаны стробилы Zamia furfuracea, а в правом нижнем углу – отдельная чешуя. На примере двух последних рисунков хорошо видна еще одна общая тенденция - сокращение числа семяпочек на мегаспорофилл: в данном случае от 4-8 у Cycas до 2 у Zamia. Источник рисунка тот же: Dörken, Rozynek, 2013.
Термин «шишка» в строгом смысле применим не ко всем голосеменным. Почему – будет сказано чуть ниже. Однако уже все роды саговниковых, кроме Cycas, имеют компактный мегастробил с жесткими чешуями. На снимке показаны стробилы Zamia furfuracea, а в правом нижнем углу – отдельная чешуя. На примере двух последних рисунков хорошо видна еще одна общая тенденция - сокращение числа семяпочек на мегаспорофилл: в данном случае от 4-8 у Cycas до 2 у Zamia. Источник рисунка тот же: Dörken, Rozynek, 2013.

Шишка в разных ее формах – это магистральная линия эволюции голосеменных. Но от нее не так уж мало уклонений на уровне подкласса, порядка, семейства, даже рода. Самое древнее – это гинкговые. Типичные мегастробилы голосеменных имеют совсем короткую ножку, а большая часть оси занята густо расположенными чешуями. У гинкго всё устроено с точностью до наоборот: ножка очень длинная и тонкая, мегаспорофиллы совсем короткие; нет не только шишки, но и вообще никаких функциональных чешуй. Такими мегастробилы гинкговых, похоже, были изначально: есть окаменелости из юрского периода. С тех пор изменилось только число семяпочек. Оно сократилось с нескольких штук до единственной у современного вида. На снимке мы видим, что в период опыления они располагаются по две на одной ножке, но развивается в норме только одна. Таким образом, здесь редукция выражена значительно сильнее, чем у саговниковых: там сокращалось лишь число семяпочек на мегаспорофилл, а здесь еще и число мегаспорофиллов на мегастробил. Ссылка на источник фотографии.
 Мегастробилы гнетовых покажем на примере эфедры. Они, как и у гинкго, редуцированы до крайности. Каждый включает единственную семяпочку. У гнетовых и зеленые листья, и спорофиллы, располагаются супротивно. Так же расположены и мегасторобилы-семяпочки: напротив друг друга. Каждая пара окружена несколькими сросшимися парами супротивных стерильных чешуй, образующих подобие околоцветника. На снимке это выглядит как некий симметричный "тюльпан" с перекрестно-супротивными лепестками. Такие собрания-тюльпаны расположены по несколько штук в пазухах обычных зеленых листьев. У двух других родов гнетовых принцип такой же, но есть некоторые особенности, которые мы рассмотрим потом. Это Ephedra foliata из Восточного Средиземноморья. Фото Gideon Pisanty. Ссылка на источник фотографии.
Мегастробилы гнетовых покажем на примере эфедры. Они, как и у гинкго, редуцированы до крайности. Каждый включает единственную семяпочку. У гнетовых и зеленые листья, и спорофиллы, располагаются супротивно. Так же расположены и мегасторобилы-семяпочки: напротив друг друга. Каждая пара окружена несколькими сросшимися парами супротивных стерильных чешуй, образующих подобие околоцветника. На снимке это выглядит как некий симметричный "тюльпан" с перекрестно-супротивными лепестками. Такие собрания-тюльпаны расположены по несколько штук в пазухах обычных зеленых листьев. У двух других родов гнетовых принцип такой же, но есть некоторые особенности, которые мы рассмотрим потом. Это Ephedra foliata из Восточного Средиземноморья. Фото Gideon Pisanty. Ссылка на источник фотографии.
Перейдем, наконец, к подклассу хвойные. Когда мы смотрим по-простому на их шишку, то кажется, что если она чем-то и отличается от мегастробила саговниковых, так только размером: те же деревянистые чешуи, те же семяпочки. Нет! Она есть результат очень длительной эволюции. Чтобы заметить это, надо взглянуть слегка вооруженным глазом на шишку в период опыления.

Вот как раз такая шишка сосны обыкновенной. Давайте посмотрим на нее внимательно. Чешуи у нее располагаются парами. В каждой паре есть толстая ярко малиновая чешуя, а под ней еще одна – тонкая, светлая, как бы уже подсыхающая. Почему так? Если толстая чешуя – это мегаспорофилл, т.е. лист, то почему он находится в пазухе тонкой чешуи, то есть другого листа? Это противоречит основополагающему ботаническому закону, согласно которому в пазухе листа не может располагаться другой лист; в пазухе листа может располагаться лишь пазушный побег. Ссылка на источник фотографии.

Так оно и есть. Тонкая – это кроющая чешуя (bract), видоизмененный лист. Толстая – семенная чешуя (ovuliferous scale - именно к ней прикреплены семяпочки, а впоследствии – семена), это видоизмененный побег. На рисунке вверху представлена схема поперечного разреза через этот комплекс (кроющая чешуя + семенная чешуя). На рисунке внизу показан такой же разрез через половинку сосновой шишки. Очевидно, что такая шишка – это отнюдь не мегастробил, а компактная совокупность многих мегастробилов, каждый из которых редуцирован до единственной семенной чешуи. Для обозначения этой своеобразной структуры одно время пытались применять термин «метастробил» («мета-» по-древнегречески «после»). Но он не особо прижился. Поэтому в дальнейшем мы будем называть её просто шишкой. Ссылка на источник рисунка.

Чтобы установить эту, казалось бы, простую истину, наука работала более 100 лет. Прорыв совершил знаменитый шведский ботаник Carl Rudolf Florin, фундаментальные труды которого вышли в середине 20-го века. Он изучил все наличные окаменелости далеких предков современных хвойных и установил, как выглядели их репродуктивные структуры. Приведем лишь два рисунка из его трудов. На них представлена реконструкция двух видов древнейших палеозойских хвойных. Оба вида имели довольно длинные вильчатые (вспомните листья гинкго!) кроющие чешуи, в пазухах которых располагались мегастробилы. Уже в то время они были сильно редуцированы (если сравнивать их, например, с мегастробилами саговниковых). На верхнем рисунке мегастробил «урезан» до единственной семяпочки, но у него еще довольно много (десятки) рудиментарных стерильных чешуй. Это настоящий пазушный побег. На втором рисунке стерильных чешуй нет, зато в пазухе каждой кроющей чешуи находятся 5-7 семяпочек на длинных мегаспорофиллах (опять же вспомним гинкго). Источник рисунка – C.R. Florin. Die Koniferen des Oberkarbons und des unteren Perms. Palaeontographica B 85: 1-729 (1938–1945).

Развитие комплекса «кроющая чешуя – семенная чешуя» у современных хвойных (нижний ряд), их мезозойских (средний ряд) и палеозойских (верхний ряд) предков по представлениям современной науки. Каждый ряд показывает последовательность событий от заложения семенной чешуи в пазухе кроющей чешуи до созревания семени. Созревание семенной чешуи показано ее цветом от светло-серого к темно-серому. Развивающиеся семяпочки и созревающие семена показаны эллипсами цветом от белого до черного. Хорошо видно, что у ранних хвойных в пазухе кроющей чешуи располагалась не семенная чешуя, а редуцированный мегастробил со многими более или менее развитыми чешуями. Причем семяпочка образовывалась на относительно позднем этапе его развития, когда он уже достиг чуть ли не окончательной величины. У современных хвойных от пазушного мегастробила осталась строго специализированная семенная чешуя. Семяпочка при этом образуется на очень ранней стадии ее развития и располагается на ее поверхности. Источник рисунка - Andrew Leslie, Juan M Losada (2019) Reproductive Ontogeny and the Evolution of Morphological Diversity in Conifers and Other Plants. Integrative and Comparative Biology. Vol. 59 (3). P. 548–558.
Голосеменные подклассы: опыление
Напомню, что перед тем, как описывать мегастробилы, мы оставили полетать в воздухе пыльцу, которая покинула микроспорангий. Действительно, у большинства голосеменных, пыльца переносится на мегастробил ветром. Но отнюдь не у всех. Так, в опылении некоторых голосеменных участвуют насекомые. Причем, это есть и в самом древнем подклассе (саговниковые), и в самом продвинутом (гнетовые). Это означает, что энтомофилия в эволюции голосеменных независимо возникла, по крайнем мере, дважды. Облигатных энтомофилов (которые опыляются только насекомыми) среди голосеменных немного, факультативных (которые опыляются и ветром, и насекомыми) гораздо больше.
У многих саговниковых довольно питательная пыльца. Кроме того, их крупные микростробилы с обширными внутренними полостями отлично подходят как спокойные места для спаривания насекомых и отложения яиц. В мегастробилах пыльцы нет, насекомым нет причин их посещать. Поэтому у некоторых видов, в опылении которых участвуют насекомые, отмечено интересное явление: мегастробилы мимикрируют под микростробилы. Насекомые по ошибке попадают в мегастробил, ничего вкусненького не находят, но доставляют туда пыльцу. У саговниковых это далеко не единственный способ привлечения насекомых. Об их разнообразии говорит хотя бы разнообразие запахов, которые издают микростробилы: от приятного (для пчел) до неприятного (может быть, для мух?).
 Чаще других в опылении саговниковых участвуют жуки-долгоносики. На снимке австралийская Macrozamia fraseri. Жуки откладывают яйца в микростробил. Молодые насекомые появляются на свет среди пыльцы, затем, переползая с растения на растение, производят опыление. На поверхности их тела есть даже специальные приспособления для переноса пыльцы. Это взаимодействие между жуками и саговниковыми документировано по окаменелостям уже в начале юрского периода, когда покрытосеменные были еще в проекте. Фото D.Blumer. Источник фотографии.
Чаще других в опылении саговниковых участвуют жуки-долгоносики. На снимке австралийская Macrozamia fraseri. Жуки откладывают яйца в микростробил. Молодые насекомые появляются на свет среди пыльцы, затем, переползая с растения на растение, производят опыление. На поверхности их тела есть даже специальные приспособления для переноса пыльцы. Это взаимодействие между жуками и саговниковыми документировано по окаменелостям уже в начале юрского периода, когда покрытосеменные были еще в проекте. Фото D.Blumer. Источник фотографии.

Для гнетовых опыление насекомыми, похоже, первичный признак. Некоторые виды потом перешли к опылению ветром. На снимке один из видов эфедры и насекомые, проявляющие интерес к его генеративным органам в период опыления. Как видите, они довольно разнообразны. На трех фотографиях в правом ряду крупным планом показаны части тела разных насекомых, буквально облепленные пыльцой. Источник рисунка - Bolinder K, Humphreys AM, Ehrlén J, Alexandersson R, Ickert-Bond SM, Rydin C (2016) From near extinction to diversification by means of a shift in pollination mechanism in the gymnosperm relict Ephedra (Ephedraceae, Gentales). Bot J Linn Soc 180: 461–477.

Опыление ветром – это тоже не так просто, как может показаться на первый взгляд. Ведь пыльца – не простая пыль, а шишка с семяпочками – не поверхность пола под шкафом. И пыльца, и шишка – это сложнейшие, очень тонко организованные произведения природы, над которыми она работала миллионы лет. Аэродинамика ветроопыления – это пусть и не большая, но самостоятельная область экологии, имеющая и свои методы исследования, и серьезные достижения. Например, установлено, что воздушный поток, несущий пыльцу, хаотичен лишь до тех пор, пока он не приблизился к шишке. Стоит ему приблизиться, как он становится упорядоченным, несущим пыльцевые зерна строго на микропиле семяпочек. Это как раз показано на рисунке, который понятен без специальных пояснений. Форма шишки довольно сложна и уникальна для каждого вида. Форма пыльцевого зерна - тоже. Они идеально подходят друг к другу, как конкретный ключ к конкретному замку. Это позволяет резко повысить эффективность опыления. Источник рисунка - Niklas, K. J. (1985) The aerodynamics of wind pollination. Botanical Review 51: 328–386.
Когда мы обсуждали репродуктивный цикл голосеменных, то отмечали, что их важнейшим достижением стала полная независимость воспроизводства от внешней капельно-жидкой воды. Так оно и есть. Но это отнюдь не означает, что вся сексуальная жизнь происходит у них «всухую». Просто они не ждут дождя и не живут в воде, как папоротники, а все необходимые половые жидкости (sexual fluids) производят сами. Конкретно тогда, когда это необходимо, и конкретно в том объеме, который нужен. Это так называемая опылительная капля, о которой мы уже упоминали вскользь, когда описывали репродуктивный цикл сосны.
 У голосеменных налицо два принципиально разных типа опылительной капли. На вид они похожи. Слева Ginkgo biloba, одиночная семяпочка; справа Juniperus communis из подкласса хвойные, шишка с 3 семяпочками. Масштабная линейка 1 мм. Источник рисунка - Nepi, M., P. von Aderkas, R. Wagner, S. Mugnaini, A. Coulter & E. Pacini. 2009. Nectar and pollination drops: how different are they? Annals of Botany 104: 205–219.
У голосеменных налицо два принципиально разных типа опылительной капли. На вид они похожи. Слева Ginkgo biloba, одиночная семяпочка; справа Juniperus communis из подкласса хвойные, шишка с 3 семяпочками. Масштабная линейка 1 мм. Источник рисунка - Nepi, M., P. von Aderkas, R. Wagner, S. Mugnaini, A. Coulter & E. Pacini. 2009. Nectar and pollination drops: how different are they? Annals of Botany 104: 205–219.
Первый тип характерен для двух древних подклассов: саговниковых и гинкговых. У них половая жидкость выделяется женским гаметофитом. Она важна не только для опыления (улавливания пыльцы), но и для оплодотворения. Это скорее оплодотворительная, чем опылительная капля: как бы переходный тип от папоротниковидных предков к более продвинутым подклассам голосеменных. Тут надо сказать, что при обсуждении репродуктивного цикла папоротников мы пропустили один важный момент. Как сперматозоид папоротника находит яйцеклетку, когда плывет к ней во время дождя? У него ведь нет времени и сил тыкаться куда попало. Он должен прицельно попасть куда следует. Оказывается, ориентироваться ему помогает слизь, которая специально образуется при плановом разрушении клеток архегония. Она частично растворяется в воде. Сперматозоид способен ощущать наличие этой слизи плыть по градиенту ее концентрации прямо к цели.
У саговниковых и гинкговых женский гаметофит также выделяет половую жидкость. Она ни с какой внешней водой не смешивается, а заполняет специальную камеру над архегониями и выпячивается из семяпочки наружу через микропиле в виде капли. Пыльца попадает на каплю? Мужской гаметофит образует пыльцевую трубку, сперматозоид выходит из нее, плывет к яйцеклетке и оплодотворяет ее.
 Опылительная капля у гинкго. (A) Формирование пыльцевой камеры перед образованием опылительной капли. (B) Разрушение клеток нуцеллуса параллельно с формированием опылительной капли. (C) Выделение опылительной капли от пыльцевой камеры в сторону микропиле. (D) Опылительная капля достигла максимального объёма к началу процесса опыления. I – интегумент, Mc – микропилярный канал, Mi – микропиле, Nu – нуцеллус (мегагаметофит), Pc – пыльцевая камера, PD – опылительная капля (pollination drop). Источник рисунка - Jin B, Jiang XX, Wang D, Zhang L, Wan YL, Wang L (2012b) The behavior of pollination drop secretion in Ginkgo biloba L. Plant Signal Behav 7:1168–1176.
Опылительная капля у гинкго. (A) Формирование пыльцевой камеры перед образованием опылительной капли. (B) Разрушение клеток нуцеллуса параллельно с формированием опылительной капли. (C) Выделение опылительной капли от пыльцевой камеры в сторону микропиле. (D) Опылительная капля достигла максимального объёма к началу процесса опыления. I – интегумент, Mc – микропилярный канал, Mi – микропиле, Nu – нуцеллус (мегагаметофит), Pc – пыльцевая камера, PD – опылительная капля (pollination drop). Источник рисунка - Jin B, Jiang XX, Wang D, Zhang L, Wan YL, Wang L (2012b) The behavior of pollination drop secretion in Ginkgo biloba L. Plant Signal Behav 7:1168–1176.
Второй тип опылительной капли характерен для гнетовых и хвойных: жидкость выделяется тканями спорофита и предназначена лишь для улавливания пыльцы. Пыльца попадает на поверхность капли, смачивается ею, набухает, тонет, опускается прямо на поверхность нуцеллуса и уже там образует пыльцевую трубку. В ней вместо сперматозоидов находятся спермии – неподвижные мужские гаметы. Подвижность им и ни к чему. Пыльцевая трубка доставляет их прямо к яйцеклетке.

Есть другой вариант, предназначенный для тех случаев, когда микропиле семяпочки обращено вниз. Против вектора гравитации пыльца тонуть в капле не может. Для этого природа придумала оригинальный механизм. Помните, когда мы обсуждали форму пыльцевых зерен, был один интересный тип: с воздушными мешками. Он характерен для некоторых родов хвойных. Обычно люди, когда видят изображение этого типа пыльцы, думают, что мешки предназначены для облегчения полета. Может быть, они и помогают пыльце летать, снижая ее удельный вес, но главное их назначение совсем не в этом. Посмотрите на картинку. Это Podocarpus neriifolius из Юго-Восточной Азии. У него семяпочки одиночные, примерно как у гинкго. Но обращены вниз. Пыльца попадает на кроющую чешую. Из микропиле выпячивается опылительная капля, которая, как любая капля, не растекается за счет поверхностного натяжения. Она распространяется до чешуи и как бы слизывает с нее пыльцу. Пыльцевые зерна мешками-поплавками вверх всплывают до нуцеллуса и там прорастают. Получается красиво и эффективно. Такой способ доставки пыльцы к нуцеллусу характерен и для многих наших хвойных. Это лишь один из опылительных механизмов. У хвойных их не менее 5. Есть пыльца без воздушных мешков, есть семяпочки без опылительной капли. Мы ознакомимся с ними в дальнейшем. Источник рисунка - Andrew B. Leslie, et al. Trait integration and macroevolutionary patterns in the pollination biology of conifers. Evolution, 2015; doi: 10.1111/evo.12670.
Опылительная капля – это не вода, а очень сложный "коктейль", каждый из компонентов которого - не лишний: минеральные и органические вещества, белки и углеводы, ферменты и гормоны. Опылительная капля не просто улавливает пыльцу и служит водной средой для мужских гамет. Она защищает весь процесс от инфекций, стимулирует мужской гаметофит к развитию в правильном направлении, причем, конкретно свой (данного вида), а не чужой. Еще одна ее функция – привлечение насекомых. Многие виды с участием насекомых в опылении четко отличаются от анемофильных (опыляемых ветром) родственников с высоким содержанием растворимых сахаров в опылительной капле. Так последняя становится еще и нектаром, вознаграждающим насекомых за участие в опылении. Получается, что нектар у голосеменных образуется совсем не так, как у цветковых. Голосеменные «изобрели» его самостоятельно.
Голосеменные подклассы: рост пыльцевой трубки, мужские гаметы
Теперь вернемся к пыльце, которая попала на семяпочку и изготовилась прорастать, т.е. давать пыльцевую трубку. Мы уже немножко касались этого вопроса. Рассмотрим его подробнее. У двух примитивных подклассов пыльцевая трубка совершенно не такая, как у двух продвинутых. У саговниковых и гинкговых налицо переходный вариант от зеленого свободноживущего мужского гаметофита папоротников к собственно пыльцевой трубке. Их пыльцевая трубка – гаусториальная. Что это значит? Вообще гаустория – это видоизмененный корень паразитических растений, предназначенный для внедрения в ткани хозяина. Получается, что нуцеллус тут как бы хозяин, а пыльцевая трубка – как бы паразит.
 На снимке показана схема этого процесса у Zamia из саговниковых. Пыльцевая трубка (Pt, на схеме их три) ведет себя очень агрессивно: внедряется в ткань нуцеллуса и даже в отдельные клетки этой ткани, использует их питательные вещества для собственного роста. В результате этой ее деятельности на месте разрушенной части нуцеллуса образуется коррозионная пыльцевая камера (С). Она заполняется опылительной каплей, в которую с противоположной от пыльцевой трубки стороны из пыльцевого зерна выходят мужские гаметы. Ссылка на источник рисунка.
На снимке показана схема этого процесса у Zamia из саговниковых. Пыльцевая трубка (Pt, на схеме их три) ведет себя очень агрессивно: внедряется в ткань нуцеллуса и даже в отдельные клетки этой ткани, использует их питательные вещества для собственного роста. В результате этой ее деятельности на месте разрушенной части нуцеллуса образуется коррозионная пыльцевая камера (С). Она заполняется опылительной каплей, в которую с противоположной от пыльцевой трубки стороны из пыльцевого зерна выходят мужские гаметы. Ссылка на источник рисунка.
{kind=link}

У гинкго пыльцевая трубка ведет себя примерно так же. Отличие такое: она не прёт в нуцеллусе напролом и не разрушает клетки, но активно ветвится в межклетниках. Она тоже питается за счет нуцеллуса, но делает это нежно, аккуратно. На рисунке весь процесс показан очень наглядно, в динамике. Ссылка на источник рисунка.

У хвойных и гнетовых (на схеме - сосна) пыльцевая трубка почти не ветвится и почти не питается за счет нуцеллуса. Она вообще не делает никаких лишних движений. Просто растет по межклетникам напрямую к архегонию и врастает в него. Мужские гаметы находятся в кончике трубки. Это характерно также и для цветковых, но клеточная организация пыльцы и пыльцевой трубки там другая. Ссылка на источник рисунка.
{kind=link}
Другие и скорости. У цветковых скорость роста пыльцевой трубки варьирует от 80 до 600 мкм/час. Голосеменные на этом фоне выглядят очень бледно: Zamia (саговниковые) -1 мкм/час; Ginkgo - 2 мкм/час; Agathis (хвойные) - 6 мкм/час; Gnetum - 5 мкм/час. Такие же огромные различия есть и в продолжительности периода от опыления до оплодотворения. У цветковых он часто измеряется немногими часами. Среди голосеменных рекордом владеет эфедра из гнетовых: примерно сутки. У остальных этот период нередко продолжается целый год. Ну что же: хвойные были первопроходцами в этом вопросе. А первопроходцы обычно не спешат.
 О мужских гаметах. С ними все хорошо знакомы на примере человеческого сперматозоида: подтянутого, элегантного и фактурного. У растений они выглядят не так ярко, а у голосеменных и вовсе не похожи ни на какие другие. Сразу же покажем вот такую картинку, где представлено всё разнообразие мужских гамет у наземных растений. 12-14 – высшие споровые со сложными, лохматыми и мохнатыми, причудливыми сперматозоидами, 1-6 – цветковые с простыми и понятными бесформенными спермиями. Всё остальное – голосеменные: очень хорошо видна их принципиальная разнородность и переходный характер. Источник рисунка - D. Southworth and M. Cresyi (1997) Comparison of Flagellated and Nonflagellated Sperm in Plants. American Journal of Botany 84(9): 1301-1311.
О мужских гаметах. С ними все хорошо знакомы на примере человеческого сперматозоида: подтянутого, элегантного и фактурного. У растений они выглядят не так ярко, а у голосеменных и вовсе не похожи ни на какие другие. Сразу же покажем вот такую картинку, где представлено всё разнообразие мужских гамет у наземных растений. 12-14 – высшие споровые со сложными, лохматыми и мохнатыми, причудливыми сперматозоидами, 1-6 – цветковые с простыми и понятными бесформенными спермиями. Всё остальное – голосеменные: очень хорошо видна их принципиальная разнородность и переходный характер. Источник рисунка - D. Southworth and M. Cresyi (1997) Comparison of Flagellated and Nonflagellated Sperm in Plants. American Journal of Botany 84(9): 1301-1311.
Так же, как опылительные капли и пыльцевые трубки, мужские гаметы голосеменных бывают двух контрастных типов: (1) сперматозоиды у саговниковых и гинкговых, (2) спермо-ядра у хвойных и гнетовых.

Сперматозоид Zamia integrifolia. Имеет форму круглодонного казанка с выпуклой крышкой. Обратите внимание на обилие ресничек, пульсация которых позволяет сперматозоиду плавать в опылительной капле. Их не много и не мало, а до 50 000 штук. Саговниковым принадлежит мировой рекорд по размеру сперматозоидов: до 400 мкм. Это примерно в 10 раз больше, чем у человека, а если брать только тело (без жгутика и ресничек), то почти в 100 раз. У гинкго сперматозоид примерно такой же по форме, но меньше размер (дл 80 мкм) и число ресничек (до 1000 шт.). Источник рисунка - Norstog, K.J. and Nicholls, T.J. (1997). The Biology of the Cycads. Cornell University Press, Ithaca.

У хвойных и гнетовых спермии похожи на спермии цветковых тем, что это спермии: у них нет органоидов движения. Но есть два отличия. (1) Они в среднем многократно крупнее (это предположительно связано с размером генома): не столько за счет длины, сколько за счет диаметра. У голосеменных диаметр лишь чуть меньше, длины, у цветковых – в 4-5 раз. (2) У большинства хвойных и гнетовых, строго говоря, вообще нет спермиев. Есть спермо-ядра. Так получается потому, что в сперматогенной клетке делится лишь ядро, а цитокинез (образование клеточной стенки) не происходит. На фотографии – спермо-ядра сосны в цитоплазме генеративной клетки. Черная линия на фотографии – не природная. Она прорисована исследователями, чтобы всё-таки показать читателю границу спермо-ядра. Спермо-ядра без клеточных стенок - очень редкое явление. С другой стороны, вполне логичное. Незнайка говорил так: «Зачем раздеваться, если завтра придется одеваться снова?». Так и тут: зачем одеваться? Это тем более актуально для сексуальной жизни: она ведь обычно происходит голышом. Fernando, D. D., Quinn, C. R., Brenner, E. D., and Owens, J. N. (2010). Male gametophyte development and evolution in extant gymnosperms. Int. J. Plant Sci. 4, 47–63.
Голосеменные подклассы: семяпочка, женские гаметы, оплодотворение
Итак, мужская гамета у нас есть. Теперь наша задача подготовить ей пару – женскую гамету. Сделаем это на примере сосны обыкновенной по рисунку из российского учебника.

Развитие семяпочки (семязачатка), как и развитие любого органа, начинается с малюсенького бугорка активно делящихся клеток. Это будущий мегаспорангий, который у голосеменных называется нуцеллусом. Когда он немножко подрастает, в его основании дифференцируется валик. Это будущий покров нуцеллуса – интегумент. Они продолжают расти параллельно, но интегумент растет чуть быстрее, поэтому постепенно обрастает нуцеллус со всех сторон. Лишь в дистальной части остается небольшое отверстие – пыльцевход или микропиле. На рисунке оно почему-то не обозначено. Нуцеллус состоит из однородной паренхимной ткани, но в его недрах обособляется одна более крупная клетка, которая называется археспориальной. В этой единственной клетке происходит мейоз (редукционное деление). Оно обозначено на рисунке розовой буквой R! и знаменует переход от диплоидного бесполого поколения (спорофита) к гаплоидному половому (гаметофиту, в данном случае, женскому). Из одной диплоидной археспориальной клетки образуется 4 гаплоидных мегаспоры. В женский гаметофит развивается лишь одна из них, остальные постепенно дегенерируют. Развитие начинается со свободноядерной стадии, когда ядра активно делятся, но остаются внутри одной большой клетки. Это на рисунке не отражено. Потом происходит целлюляризация: образование клеточных стенок. Потом клетки (их много: тысячи) дорастают до окончательного размера. Так формируется гаплоидный женский гаметофит, который в семени будет называться эндоспермом и использоваться для питания зародыша и проростка. Параллельно формируются архегонии (два или несколько). Архегонии устроены просто: несколько мелких малозначимых клеток + единственная огромная яйцеклетка. Ссылка на источник рисунка.
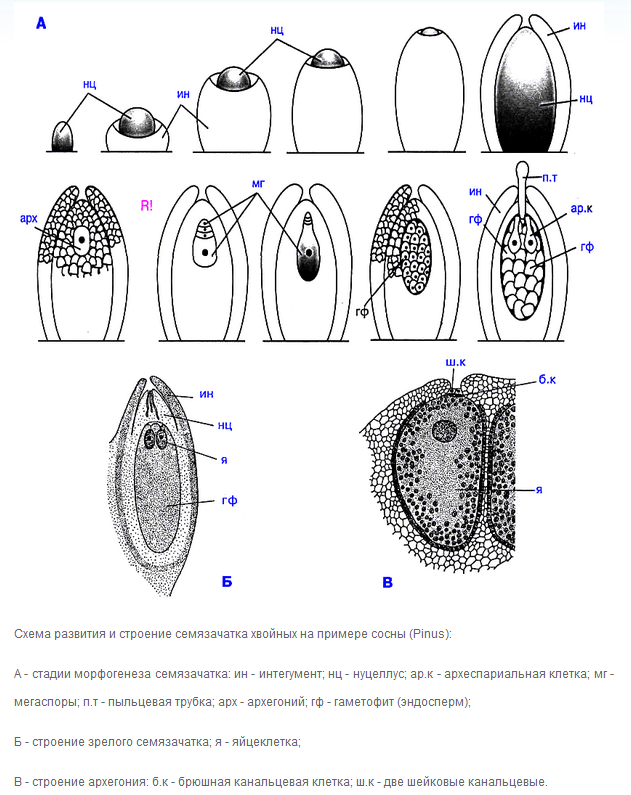{kind=link}
Это лишь один, типичный, современный вариант развития. Однако голосеменные живут на нашей планете давно, причем, всё самое интересное в их эволюции произошло ещё в палеозое. Приведем лишь один пример.
 У первых палеозойских семенных папоротников семяпочки были без интегумента, а семена – без кожуры. Покровы, конечно, были, но не сплошные и не сросшиеся с внутренними частями семяпочки. Эволюция (ряд слева направо) шла понятно в какую сторону. У последнего варианта в этом ряду процесс завершился образованием настоящего сплошного интегумента. Источник рисунка - Рейвн П., Эверт Р., Айкхорн С. Современная ботаника: В 2-х т. Пер. с англ. - М.: Мир, 1990.
У первых палеозойских семенных папоротников семяпочки были без интегумента, а семена – без кожуры. Покровы, конечно, были, но не сплошные и не сросшиеся с внутренними частями семяпочки. Эволюция (ряд слева направо) шла понятно в какую сторону. У последнего варианта в этом ряду процесс завершился образованием настоящего сплошного интегумента. Источник рисунка - Рейвн П., Эверт Р., Айкхорн С. Современная ботаника: В 2-х т. Пер. с англ. - М.: Мир, 1990.
Четыре современных подкласса обособились тоже давным-давно. Есть ли между ними какие-то различия в части структуры и развития семяпочек. Есть. Причем, не только между, но и внутри. Они подробно описаны учеными. Однако в мелкие анатомические и цитологические детали мы пока вникать не будем. Нарисуем картину крупными мазками.
Один из общих принципов эволюции такой: развитие от большего и неопределенного числа каких-то морфологических элементов (клеток, органов) к меньшему и определенному. Возьмем, например, число архегониев на женский гаметофит. У саговниковых материнских клеток архегониев закладывается много, да и развитых архегониев бывает до 10 шт. У более продвинутых подклассов, начиная от гинкго, их обычно всего два.
Второе направление – сокращение размера микро- и макроструктур. Вот выше мы удивили вас размером сперматозоидов у саговниковых. Но в сексуальной сфере актуальна гармония и размер тут не может не иметь значения. Согласитесь, для саговниковых как-то несерьезно было бы соваться с большими сперматозоидами к маленьким яйцеклеткам. Действительно, у них и размер яйцеклетки вызывает всяческое уважение: до 6 мм (кубинский Microcycas). Необычайно велико и ядро яйцеклетки: до 5000 мкм. По сравнению с яйцеклеткой страуса (20 см), не впечатляет. Но для растений это абсолютный рекорд. У гинкго яйцеклетки меньше - до 1000 мкм, у хвойных еще меньше – около 500 мкм, у цветковых просто микроскопические: от 15 до 25 мкм.
Скорость процессов многократно возрастает от примитивных подклассов к продвинутым. Саговниковые живут в тропиках, где тепла – хоть отбавляй. Однако от появления мегаспоры до оплодотворения у них проходит от 190 до 350 дней. Гинкго живет в субтропиках, где тепла меньше. Но ему оказывается достаточно 150 дней. Наконец, многие виды хвойных и гнетовых (эфедра) спокойно проходят этот путь за месяц с небольшим даже в бореальной зоне с ее минимальной теплообеспеченностью.
 От примитивных классов к продвинутым усиливается также привязка развития интегумента, нуцеллуса и эндосперма к фактам опыления и оплодотворения. На этом рисунке показан размер семяпочек в момент опыления (pollination) и оплодотворения (fertilization) относительно размера зрелого семени (seed maturity), у разных групп семенных растений. Интегумент + нуцеллус показаны черным цветом, мегагаметофит (эндосперм) – белым, зародыш – опять же черным. Callospermarion – это семенной папоротник из позднего Карбона. Саговниковые – это Zamia + Encephalartos. Amborella – вечнозеленое деревце из Новой Каледонии, самое примитивное из современных цветковых (бессосудистое и т.д.), единственный представитель семейства, которое отделилось от основного ствола цветковых примерно 130 млн. лет назад и с тех пор мало изменилось.
От примитивных классов к продвинутым усиливается также привязка развития интегумента, нуцеллуса и эндосперма к фактам опыления и оплодотворения. На этом рисунке показан размер семяпочек в момент опыления (pollination) и оплодотворения (fertilization) относительно размера зрелого семени (seed maturity), у разных групп семенных растений. Интегумент + нуцеллус показаны черным цветом, мегагаметофит (эндосперм) – белым, зародыш – опять же черным. Callospermarion – это семенной папоротник из позднего Карбона. Саговниковые – это Zamia + Encephalartos. Amborella – вечнозеленое деревце из Новой Каледонии, самое примитивное из современных цветковых (бессосудистое и т.д.), единственный представитель семейства, которое отделилось от основного ствола цветковых примерно 130 млн. лет назад и с тех пор мало изменилось.
Что мы видим? В двух древних примитивных группах семяпочки к периоду опыления не просто достигают уже значительного размера, но и имеют внутри активно развивающийся мегагаметофит. Это ну очень расточительно: пыльцы еще нет, а вещество и энергия уже израсходованы. У гинкго и хвойных опыление происходит очень рано, когда семяпочки маленькие и незрелые, т.е. примерно на том же уровне развития, что и у цветковых. Это уже значительно эффективнее.
У саговниковых и гинкго отсутствие пыльцы не исключает дальнейший рост нуцеллуса и дифференциацию мегагаметофита (семяпочки продолжают развиваться независимо от наличия пыльцы); у хвойных исключает, это дает им преимущество.
Опыление – это только часть дела, притом, меньшая. Главная его часть, конечно, оплодотворение. Для оплодотворения голосеменным нужен развитый мегагаметофит с архегониями и достаточно жестким интегументом (иначе негде будет оформляться и функционировать пыльцевой капле). Поэтому к моменту оплодотворения мегагаметофит у всех голосеменных сильно увеличивается в размере. Это неэффективно: зародыш то ли будет, то ли нет, а рост происходит по полной программе. У современных хвойных эндосперм всё-таки заметно подрастает после успешного оплодотворения, после не успешного - дегенерирует. Идеальное завершение этой логической цепи мы видим у цветковых. Там эндосперм и покровы образуются совершенно по-другому, но их формирование начинается только после появления реальной здоровой зиготы. Это максимально эффективный вариант. Он возник не раньше, чем 200 млн. лет назад. До этого экономии ресурсов почти не было.
Среди голосеменных это есть лишь в самом молодом (те же 200 млн. лет) подклассе гнетовые, причем, лишь у одного рода Gnetum. Однако принцип тут совсем другой. Это отдельная, притом, очень интересная история. Здесь мы ее пропустим, а вернемся к ней тогда, когда дело дойдет до специального раздела о гнетовых. Гнетовые, как вы заметили, вообще любят «выпендриться» не только перед остальными группами растений, но и друг перед другом. А способ оплодотворения – это, пожалуй, самая любимая ими область эволюционного творчества. Повторюсь, мы вернемся к этому в свое время и в своем месте. Источник рисунка - Andrew B. Leslie and C. Kevin Boyce. 2012. Ovule Function and the Evolution of Angiosperm Reproductive Innovations. International Journal of Plant Sciences. Vol. 173, N. 6. P. 640-648.
На этом же рисунке серым цветом в правом вертикальном ряду показан абсолютный размер зрелых семян. Эти данные не вполне корректно характеризуют реальную ситуацию. Так, у семенного папоротника Callospermarion семя маленькое. Но были и совсем другие семенные папоротники с семенем длиной до 7 см (например, Pachytesta gigantea из позднего Карбона). Гнетум также вряд ли характеризует гнетовые в целом. Например, у эфедры семена, в основном, мелкие, примерно как у хвойных. Но если исключить эти два неудачных примера, эволюционная тенденция очевидна: от примитивных подклассов к продвинутым размер семяпочки и семени убывает. Понятно почему: ведь голосеменные – это переходное звено от высших споровых с их свободноживущим женским гаметофитом к цветковым, у которых женский гаметофит – это всего 8 клеток.

Покажем еще одну картинку: длина и диаметр семяпочек в момент опыления (A), мегагаметофита в момент оплодотворения (B) у 6 видов вымерших голосеменных (белые треугольники), 36 видов современных голосеменных (белые кружки) и 84 видов цветковых (черные кружки). Обратите внимание, что шкала на обеих осях не простая, а логарифмическая. Мы видим, что вымершие голосеменные в части размера семяпочек и мегагаметофита не отличаются от современных. Это понятно: ведь базовый принцип организации у них один и тот же. А вот от цветковых голосеменные отличаются существенно.
По размеру семяпочек в период опыления различия меньше. Семяпочки в период опыления в обоих случаях маленькие. Но… по разным причинам: у цветковых они в принципе маленькие, а у голосеменных просто находятся на очень ранней стадии развития. Тем не менее, различия существенны. Средняя длина у голосеменных 1730, у цветковых 480 мкм. Понятно почему: у цветковых семяпочка имеет меньше функций: она не взаимодействует с пыльцевым зерном, не отвечает за опылительную каплю и т.д. Налицо огромное разнообразие внутри сравниваемых групп: диапазоны разнообразия перекрываются почти наполовину. Так, у Metasequoia из хвойных длина семяпочки всего 200–250 мкм. Однако даже самые мелкие голосеменные семяпочки всё-таки значительно крупнее, чем самые мелкие семяпочки цветковых (в некоторых семействах меньше 100 мкм).
По размеру мегагаметофита в момент оплодотворения различия просто гигантские, на несколько порядков, причем, диапазоны разнообразия не перекрываются вообще. Причину мы уже обсуждали: у голосеменных мегагаметофит – это, в основном, будущий эндосперм, ткань для питания зародыша. У цветковых эндосперм образуется совсем по-другому и в другое время. Источник рисунка тот же – Leslie, Boyce, 2012.
Голосеменные подклассы: семена
Размер семян – важный признак в экологии растений. Перед эволюцией всегда есть выбор: много мелких или мало крупных. В каждом случае она решает этот вопрос с учетом множества разнообразных факторов, образующих сложный клубок. Диапазон размера семян – 10 порядков: от 0,0001 мг у некоторых орхидей до 42 кг у двойных кокосов (сейшельской пальмы). Как видите, оба края этого диапазона относятся к цветковым. У голосеменных диапазон значительно меньше: 5 порядков. Нет очень мелких и очень крупных семян.
Главный фактор, определяющий размер семян – это способ их распространения. Главная закономерность простая. Семена, распространяемые ветром, мельче, они имеют для этого специальные приспособления. У голосеменных это обычно разного рода и формы кожистые крылья. Семена, распространяемые животными крупнее; многие из них имеют еще и специальные питательные ткани для привлечения потребителей.
 В экологии растений есть такой термин – «пропагула» (от латинского глагола «propago» - размножать, распространять). Пропагула – это часть растения, которая отделяется от него с целью размножения (расселения). Пропагулы – это и разнообразные органы вегетативного размножения, и споры у споровых растений, и целые плоды, и отдельные семена. В верхней части этого рисунка показаны 5 типов пропагул у голосеменных. Обычным, неспециализированным (Unspecialized) типом у голосеменных считаются семена с крылом, предназначенные для распространения ветром.
В экологии растений есть такой термин – «пропагула» (от латинского глагола «propago» - размножать, распространять). Пропагула – это часть растения, которая отделяется от него с целью размножения (расселения). Пропагулы – это и разнообразные органы вегетативного размножения, и споры у споровых растений, и целые плоды, и отдельные семена. В верхней части этого рисунка показаны 5 типов пропагул у голосеменных. Обычным, неспециализированным (Unspecialized) типом у голосеменных считаются семена с крылом, предназначенные для распространения ветром.
Все остальные типы – специализированные (Specialized) для распространения животными. Seed – это просто крупные семена без крыльев. Их эндосперм привлекает животных как продукт питания. Значит, для размножения эти семена хороши лишь в том случае, если животные не съедают все семена, а часть прячут в почву, они потом прорастают. Остальные типы пропагул обозначены словом «Fleshy» - мясистый, англ. Тут возможны три варианта: ‘Fleshy I’ – срослись и стали мясистыми чешуи шишки («шишкоягоды»), ‘Fleshy II’ – мясистые кроющие чешуи или что-то еще с частичным покрытием единственного семени, ‘Fleshy III’ – единственное семя полностью покрыто мясистой тканью. Во всех этих трех типах животными усваивается лишь питательный «околоплодник». Сами семена проходят через пищеварительный тракт неповрежденными. Более того, их всхожесть от этого часто повышается. На диаграммах показано разнообразие семян по объему (куб. мм) у 4 современных голосеменных подклассов по типам пропагул. Шкала объема семян – логарифмическая. Линиями показан сплошной диапазон разнообразия, кружками – отдельные виды за пределами сплошного диапазона.
Как видите, в двух древних примитивных подклассах (саговниковые и гинкговые) все семена одного типа: максимально крупные, покрытые мясистой питательной тканью. Есть такие семена и у гнетовых. В этом подклассе появляются также пропагулы с мелкими семенами, которые частично прикрыты мясистыми тканями. Наконец, у хвойных наблюдаются все возможные типы пропагул. При этом максимальный размер имеют крупные семена, привлекающие животных питательным эндоспермом, а также семена, полностью покрытые мясистой таканью. Минимальный размер имеют неспециализированные семена, распространяемые ветром, и семена из «шишкоягод». Источник рисунка - Leslie, A.B., Beaulieu, J.M., Mathews, S. 2017.Variationin seed size is structured by dispersal syndrome and cone morphology in conifers and other non-flowering seed plants. New Phytologist. DOI: 10.1111/nph.14456.
Каждая из этих адаптаций к распространению семян возникала в ходе эволюции голосеменных не один и не два раза. Поэтому конкретные способы формирования тканей с похожей функцией в разных филогенетических линиях разные. Старые голосеменные подклассы возникли и развивались в эпоху динозавров. Динозавры любили полакомиться их семенами. Будучи животными серьезного размера, мелкими семенами они заинтересовались бы вряд ли. Поэтому семена формировались довольно крупными. У современных саговниковых они в среднем имеют длину 3-4 при толщине 2-3 см. Кстати, слоны в Африке до сих пор считают семена некоторых энцефаляртосов деликатесом, активно ищут их и поедают. Для выживания древним голосеменным надо было устроить дело так, чтобы животные распространяли семена, но не портили их. У цветковых этой задаче идеально отвечает плод костянка: сочный и питательный снаружи, твердый и надежный внутри. Вопрос решается через 3-слойный околоплодник (например, у косточковых плодовых деревьев): тонкий кожистый внеплодник, мясистый межплодник и одревесневший внутриплодник. У голосеменных нет плодов, которыми можно было бы привлечь животных. Пришлось привлекать их самими семенами. Интегумент не так прост, как кажется спервоначалу. В полном варианте он состоит из трех слоев. Наружный слой семенной кожуры (саркотеста) мягкий и мясистый (то же у граната), средний слой (склеротеста) твёрдый, а внутренний слой (эндотеста) к моменту созревания семени плёнчатый. Животные заглатывают семя целиком и переваривают саркотесту, не повреждая склеротесты. На тот случай, если они окажутся чересчур зубастыми и захотят разгрызть склеротесту, эндосперм многих саговниковых содержит токсины. Поэтому большинство потребителей ограничиваются мякотью саркотесты, на эндосперм и зародыш не посягают.
 Семена гинкго. Хорошо видны все три «тесты». У гинкго семена опадают на землю и издают очень неприятный запах. Он отпугивает белочек, бурундучков, мышей и других потребителей ореха. Понятно зачем: растение «заинтересовано» не в том, чтобы его семена разгрызали, а в том, чтобы их глотали целиком. Так и действуют хищники привлеченные запахом гниющего мяса. Здорово «придумано»! Ссылка на источник фотографии.
Семена гинкго. Хорошо видны все три «тесты». У гинкго семена опадают на землю и издают очень неприятный запах. Он отпугивает белочек, бурундучков, мышей и других потребителей ореха. Понятно зачем: растение «заинтересовано» не в том, чтобы его семена разгрызали, а в том, чтобы их глотали целиком. Так и действуют хищники привлеченные запахом гниющего мяса. Здорово «придумано»! Ссылка на источник фотографии.
{kind=link}

Развитая мясистая саркотеста есть также у некоторых гнетовых и хвойных. Однако в большинстве случаев интегумент к моменту зрелости становится обычной деревянистой семенной кожурой. Правда, у некоторых хвойных важное значение для распространения семян саркотеста приобретает совсем другим способом: из нее формируется кожистое крыло, помогающее распространению семян ветром. На снимке семена туи западной (Thuja occidentalis). Ссылка на источник фотографии.

У других хвойных крыло с точно такой же функцией образуется сосем по-другому: не из интегумента семени, а из семенной чешуи. Это все наши хвойные: сосна, ель, пихта, лиственница. На снимке семенная чешуя гималайской Pinus wallichiana. Хорошо видно, что крыло происходит не от интегумента, а именно от семенной чешуи. Фото Jan De Laet. Ссылка на источник фотографии.

В продвинутых подклассах (гнетовые и хвойные) тоже немало родов и видов, которые привлекают насекомых мясистыми и питательными тканями, окружающими семена. На снимках Ephedra aphylla из Палестины (слева вверху, фото Avinoam Danin), Halocarpus bidwilli из Новой Зеландии (справа вверху), Taxus baccata (слева внизу) и Juniperus communis (справа внизу, фото Василия Гелюты) из Европы. Все эти адаптации возникли независимо одна от другой, на совершенно разной морфологической основе. Объединяет их размер пропагул и отдельных семян: они мелкие, спокойно помещаются в ротовую полость и проходят через пищевод небольшой птички или мышки. Оно и понятно: динозавры давно и окончательно приказали долго жить. Их место заняли значительно более разнообразные по размеру млекопитающие и птицы. На них теперь и ориентируется большая часть современных голосеменных. Источики фотографий: слева вверху, справа вверху, слева внизу, справа внизу.

Крупные семена нового типа, без мясистых тканей и с относительно мягкой кожурой появились у голосеменных не так давно. Они имеют смысл лишь в том случае, если им соответствуют виды животных, способные прятать такие семена в почве и запоминать, где они спрятаны. Семена этого типа стали появляться лишь после того, как млекопитающие и особенно птицы прошли большой цикл прогрессивной эволюции мозга и высшей нервной деятельности. В настоящее время к такому взаимодействию с растениями способны почти исключительно представители интеллектуальной элиты пернатого сообщества: семейство врановые (Corvidae). Крупные бескрылые семена есть лишь в подклассе хвойные, но не только у хорошо известных нам кедровых сосен, но и, например, у араукарии. На снимке австралийская Araucaria bidwillii. Для врановых ее семя крупновато (17,5 г). Поэтому тут задействованы большие попугаи, у которых с мозгами тоже всё в порядке. Ссылка на источник фотографии.
Голосеменные подклассы: зародыш и проросток
 О развитии зародыша. Начнем с древнейшего варианта – саговниковых (на примере Cycas). Первое деление зиготы (oospora) превращает ее в проэмбрио (предзародыш). Интересно, что как и при делении мегаспоры, делятся сначала лишь свободные ядра (free-nuclei), а клеточные стенки не образуются. Первые клеточные деления обычно синхронные. Число свободных ядер очень большое, до 1000 шт. Потом клеточные стенки всё-таки начинают понемножку образовываться. Следом начинается дифференциация клеток. В это время проэмбрио состоит из трех частей (от микропилярного полюса к халазальному): зона свободных ядер, зона так называемого подвеска (суспензора = suspensor cells) и зона собственно зародышевых инициальных клеток. На следующем этапе клетки подвеска начинают активно удлиняться. Задача подвеска – как бы прорасти через женский гаметофит и протолкнуть перед собой инициальные клетки зародыша (embryonal cells) в центральную часть мегегаметофита (будущего эндосперма семени). Подвесок отчасти напоминает кончик корня: как корешок среди частичек почвы, он прокладывает себе дорогу среди клеток женского гаметофита, частично растворяя их при помощи собственных ферментов и используя их содержимое для питания. От этого в центре женского гаметофита формируется так называемая коррозионная полость, в которой потом будет развиваться зародыш. Подвески у саговниковых большие, рекордные по размеру (до 7 см). Когда зародышевые инициали оказываются достаточно продвинутыми в мегагаметофит, рост подвеска прекращается, а проэмбрио начинает дифференцироваться на органы и ткани, т.е. превращается в собственно зародыш (эмбрио). Подвесок содействует питанию зародыша. Зародыш развивается в коррозионной полости и постепенно заполняет ее. У зрелого зародыша имеются зародышевый корень, гипокотиль (подсемядольное колено), зародышевая почка (плюмула) и семядоли (2 и более, до 18), которые окружают плюмулу. Подвесок и свободноядерная зона по мере развития зародыша постепенно дегенерируют. Ссылка на источник рисунка.
О развитии зародыша. Начнем с древнейшего варианта – саговниковых (на примере Cycas). Первое деление зиготы (oospora) превращает ее в проэмбрио (предзародыш). Интересно, что как и при делении мегаспоры, делятся сначала лишь свободные ядра (free-nuclei), а клеточные стенки не образуются. Первые клеточные деления обычно синхронные. Число свободных ядер очень большое, до 1000 шт. Потом клеточные стенки всё-таки начинают понемножку образовываться. Следом начинается дифференциация клеток. В это время проэмбрио состоит из трех частей (от микропилярного полюса к халазальному): зона свободных ядер, зона так называемого подвеска (суспензора = suspensor cells) и зона собственно зародышевых инициальных клеток. На следующем этапе клетки подвеска начинают активно удлиняться. Задача подвеска – как бы прорасти через женский гаметофит и протолкнуть перед собой инициальные клетки зародыша (embryonal cells) в центральную часть мегегаметофита (будущего эндосперма семени). Подвесок отчасти напоминает кончик корня: как корешок среди частичек почвы, он прокладывает себе дорогу среди клеток женского гаметофита, частично растворяя их при помощи собственных ферментов и используя их содержимое для питания. От этого в центре женского гаметофита формируется так называемая коррозионная полость, в которой потом будет развиваться зародыш. Подвески у саговниковых большие, рекордные по размеру (до 7 см). Когда зародышевые инициали оказываются достаточно продвинутыми в мегагаметофит, рост подвеска прекращается, а проэмбрио начинает дифференцироваться на органы и ткани, т.е. превращается в собственно зародыш (эмбрио). Подвесок содействует питанию зародыша. Зародыш развивается в коррозионной полости и постепенно заполняет ее. У зрелого зародыша имеются зародышевый корень, гипокотиль (подсемядольное колено), зародышевая почка (плюмула) и семядоли (2 и более, до 18), которые окружают плюмулу. Подвесок и свободноядерная зона по мере развития зародыша постепенно дегенерируют. Ссылка на источник рисунка.
{kind=link}
 У гинкго эмбриогенез очень похож на таковой у саговниковых. Там у нас была схема, здесь покажем фотографию. Это начало дифференциации зародыша, когда подвесок (suspensor) еще хорошо виден и продолжает функционировать. Cotyledon primordium – это зачатки семядолей. Shoot apex – зародышевая почка (плюмула). Ссылка на источник рисунка.
У гинкго эмбриогенез очень похож на таковой у саговниковых. Там у нас была схема, здесь покажем фотографию. Это начало дифференциации зародыша, когда подвесок (suspensor) еще хорошо виден и продолжает функционировать. Cotyledon primordium – это зачатки семядолей. Shoot apex – зародышевая почка (плюмула). Ссылка на источник рисунка.

У гнетовых (на примере эфедры) резко (обычно до 8 шт.) сокращено число свободных ядер в начале развития проэмбрио. Вокруг этих ядер довольно быстро формируются клеточные стенки. Каждая из 8 клеток развивается самостоятельно: делится на инициальную клетку подвеска и инициальную клетку зародыша. Подвесок растет, проэмбрио делится, потом дифференцируется в эмбрио. Между несколькими парами «подвесок - зародыш» происходит некоторое соревнование, в результате которого побеждает сильнейший: в семени обычно бывает лишь один зародыш. Ссылка на источник рисунка.
{kind=link}

У хвойных (на примере сосны) первые 1-2 деления зиготы обычно бывают не поперек, а вдоль будущего зародыша. Так получается 2 или 4 свободных ядра. Они собираются в нижней части проэмбрио и продолжают делиться уже поперек. Число свободных ядер обычно не больше 8. Потом образуются клеточные стенки. Те, кто следит за рассказом по рисунку, обратите внимание, что художник изобразил дальнейшие этапы развития не по порядку. Поэтому смотрите последовательность изображений в соответствии с последовательностью букв в латинском алфавите: H – I – J – K – L. Сначала в каждой линии появляются инициаль подвески и инициаль собственно зародыша. Подвесок сразу же развивается бурно: клетки быстро растут по направлению к основанию женского гаметофита и вырастают очень длинными. Зародышевые инициальные клетки находятся на конце растущего подвеска и в первое время не особенно активны. Первичный подвесок - это несколько параллельных продольных рядов клеток. Через некоторое время первичный подвесок трансформируется во вторичный: параллельные ряды клеток разъединяются, искривляются и дальше растут не вполне параллельно друг другу. Зародышевые инициали находятся на конце каждого ряда клеток подвеска. В процессе роста ряды как бы конкурируют между собой, в результате чего остается один из них. Подвесок ко времени полного развития зародыша дегенерирует. Ссылка на источник рисунка.
{kind=link}
Таким образом, в эволюции зародыша у голосеменных налицо следующие тенденции: резкое сокращение свободноядерной стадии (и продолжительности, и числа свободных ядер), резкое сокращение числа инициальных клеток подвеска и зародыша, появление конкуренции между парами «подвесок - зародыш» и появление сложного 2-уровневого подвеска у хвойных. Всё это повысило эффективность семенного размножения. У цветковых эти тенденции, в основном, продолжились. Так, свободноядерная стадия в развитии проэмбрио исчезла вовсе. Но это уже не наша тема. Я боюсь, что уже и без того утомил вас до крайности разными, может быть, излишними подробностями.

В эволюции всей системы семенного размножения у голосеменных также наблюдаются важные закономерности. Одна из них отражена вот на этом рисунке: рост доли зародыша от общей массы семени. У ископаемых голосеменных зародыши были маленькие. У сохранившихся до наших дней доля зародыша увеличивается от древних филогенетических групп к более современным: от 20% у саговниковых и гинкговых до 50% в молодых линиях хвойных. Кстати, точно такая же тенденция есть и у цветковых. Источник рисунка - Forbis TA, Floyd SK, deQueiroz A. 2002. The evolution of embryo size in angiosperms and other seed plants: implications for the evolution of seed dormancy. Evolution 56: 2112–2125.
У древних примитивных голосеменных подклассов (саговниковые и гинкговые) семена покидают материнское растение задолго до окончательной дифференциации зародыша, а иногда даже и до оплодотворения! Всё это происходит в отделившемся от материнского растения семени и очень медленно. Собственно говоря, у них вообще непонятно, где кончается эмбриональный этап и где начинается постэмбриональный. Семя лежит на земле без видимых изменений, т.е. формально находится в состоянии покоя, который может продолжаться до двух лет. Такой покой называется морфологическим. Зародыш развивается внутри семени и, развившись до определенного морфологического состояния, сразу же прорастает. Очевидно, что этот тип развития более или менее приемлем лишь в безморозном и равномерно-влажном климате. Ведь в любом другом климате прорастание семени в неподходящий сезон будет означать гарантированную гибель проростка. Чтобы освоить регионы с другими типами климата, голосеменным надо было серьезно поработать над своим семенным размножением. Они это сделали.
Если проводить аналогию с животными, то подклассы с недоразвитым зародышем можно соотнести с яйцекладущими. Тогда остальные семенные растения будут ближе живородящим. У гнетовых и хвойных в покидающих материнское дерево семенах зародыш полностью или почти полностью сформирован. Но это отнюдь не означает, что семя готово к немедленному прорастанию. Изредка встречается и такое. Но только там, где прорастание без периода покоя выгодно виду. Во всех остальных случаях оно происходит конкретно в оптимальное время года, после получения совершенно определенного объема совершенно определенных условий тепла и влажности, сугубо индивидуальных не только для каждого вида, но иногда даже и для каждого географического экотипа. До этого момента семена находятся в состоянии физиологического покоя. Способность к такой тонкой настройке появилась не сразу; в относительно древних филогенетических линиях встречаются разные промежуточные типы взаимодействия между семенами и климатом: морфофизиологический покой. Громадное разнообразие вариантов обеспечило гнетовым и хвойным такое же разнообразие природных условий, в которых они могут эффективно жить и размножаться.
 Собственно прорастание тоже происходит по-разному в двух группах парах подклассов. У саговниковых (на рисунке – Cycas) и гинкговых семядоли остаются внутри энодосперма. Поэтому не зеленеют, в фотосинтезе не участвуют и служат лишь для транзита питательных веществ из эндосперма семени в проросток. Ссылка на источник рисунка.
Собственно прорастание тоже происходит по-разному в двух группах парах подклассов. У саговниковых (на рисунке – Cycas) и гинкговых семядоли остаются внутри энодосперма. Поэтому не зеленеют, в фотосинтезе не участвуют и служат лишь для транзита питательных веществ из эндосперма семени в проросток. Ссылка на источник рисунка.
{kind=link}

У гнетовых и хвойных (на рисунке - Pinus) семядоли, как правило, выносят на поверхность почвы всё семя, сами постепенно освобождаются от эндосперма, оставаясь, впрочем, связующим звеном между ним и проростком до тех, пор, пока это необходимо. При этом они быстро зеленеют и активно участвуют в фотосинтезе. Ссылка на источник рисунка.
{kind=link}
Заключение
Тут можно было бы продолжить, например, обсудить значение голосеменных в народном хозяйстве. Однако я не буду этого делать. Во-первых, я и так уже утомил вас чересчур длинным повествованием. Во-вторых, это лучше перенести в другие, более конкретные разделы. Лучше закончим тем, что обсудим организацию этих самых разделов.
Три из четырех голосеменных подклассов не имеют большого значения для России и ее декоративного садоводства. Поэтому саговниковые, гинкговые и гнетовые представлены на нашем сайте одной страницей каждый. Весь остальной огромный объем информации относится к подклассу хвойные.
Подклассы делятся на порядки, порядки на семейства. Например, у покрытосеменных сотни семейств объединены в десятки порядков. У хвойных число порядков, в основном, совпадает с числом семейств. Поэтому в нашей рубрикации порядки пропущены, а подкласс хвойные делится сразу на семейства.
Каждое из тех семейств, которые не имеют большого значения для России – араукариевые (Araucariaceae), головчатотисовые (Cepholotaxaceae), подокарповые (Podocarpaceae) и сциадопитисовые (Sciadopitiaceae) – представлено у нас единственной таксономической рубрикой. В остальных трех семействах, важных для России, выделены один (тисовые, Taxaceae) или несколько (кипарисовые, Cupressaceae; сосновые, Pinaceae) родов.
Этот же принцип использован и для родов. Малозначимые для России (например, цедрус, Cedrus) или монотипые (содержащие единственный вид, например, микробиота, Microbiota) представлены единственной рубрикой. Важные роды подразделены на несколько или много видовых рубрик. При этом важные виды поименованы отдельно, а для не особенно важных есть рубрика «Другие».
Автор С.Н.Горошкевич
- Подкласс САГОВНИКОВЫЕ (Cycadeae)
- Подкласс ГИНКГОВЫЕ (Ginkgoidae)
- Подкласс ГНЕТОВЫЕ (Gnetidae)
- Pinidae - Хвойные +
- Araucariaceae - Араукариевые +
- Cepholotaxaceae - Головчатотисовые
- Cupressaceae - Кипарисовые +
- Athrotaxis - Атротаксис
- Callitris (каллитрис) и его родственники
- Callitropsis, Xanthocyparis - Каллитропсис, ксантоципарис
- Calocedrus - Калоцедрус
- Cunninghamia - Куннингамия
- Cupressus - Кипарис
- Chamaecyparis - Кипарисовик +
- Fokienia - Фокиения
- Hesperosyparis - Хеспероципарис
- Juniperus - Можжевельник +
- Juniperus chinensis - Можжевельник китайский, Juniperus procumbens - Можжевельник лежачий или распростертый
- Juniperus communis - Можжевельник обыкновенный
- Juniperus horizontalis - Можжевельник горизонтальный
- Juniperus rigida - Можжевельник твердый
- Juniperus x pfitzeriana - Можжевельник пфитцера
- Juniperus pseudosabina - Можжевельник ложноказацкий
- Juniperus sabina - Можжевельник казацкий
- Juniperus scopilirum - Можжевельник скальный
- Juniperus semiglobosa - Можжевельник полушаровидный
- Juniperus squamata - Можжевельник чешуйчатый, Juniperus pingii - Можжевельник пинга
- Juniperus virginiana - Можжевельник виргинский
- Juniperus. Другие виды
- Juniperus. Другие гибриды
- Juniperus chinensis - Можжевельник китайский, Juniperus procumbens - Можжевельник лежачий или распростертый
- Microbiota - Микробиота
- Platycladus - Платикладус
- Sequoia, Sequoiadendron, Metasequoia - Секвойя, секвойядендрон, метасквойя
- Taiwania - Тайвания
- Taxodium, Glyptostrobus, Cryptomeria - Таксодиум, глиптостробус, криптомерия
- Tetraclinis - Тетраклинис
- Thuja - Туя +
- Thujopsis - Туевик
- Athrotaxis - Атротаксис
- Pinaceae - Сосновые +
- Abies +
- Cedrus
- Larix +
- Picea +
- Picea abies
- Picea asperata
- Picea breweriana
- Picea crassifolia
- Picea engelmannii
- Picea glauca
- Picea glehnii
- Picea jezoensis
- Picea koraiensis
- Picea koyamae
- Picea mariana
- Picea meyeri
- Picea obovata
- Picea omorica
- Picea orientalis
- Picea pungens
- Picea rubens
- Picea schrenkiana
- Picea sitchensis
- Picea. Другие виды
- Picea. Гибриды
- Picea abies
- Pinus +
- Pinus albicaulis
- Pinus aristata
- Pinus banksiana
- Pinus cembra
- Pinus contorta
- Pinus densiflora
- Pinus flexilis
- Pinus heldreichii
- Pinus koraiensis
- Pinus monticola
- Pinus mugo
- Pinus nigra
- Pinus parviflora
- Pinus peuce
- Pinus ponderosa
- Pinus pumila
- Pinus rigida
- Pinus resinosa
- Pinus sibirica
- Pinus strobus
- Pinus sylvestris
- Pinus tabuliformis
- Pinus wallichiana
- Pinus. Другие виды
- Pinus. Гибриды
- Pinus albicaulis
- Pseudotsuga +
- Tsuga +
- Pinaceae. Другие роды
- Podocarpaceae - Подокарповые +
- Sciadopitiaceae - Сциадопитисовые
- Taxaceae - Тисовые +
Thuja occidentalis - Туя ...
Статья магаданских исслед...
Обзорная статья дальневос...
База данных "Хвойные...
Подкласс САГОВНИКОВЫЕ (Cy...
технология
Статья С.Н.Горошкевича “С...
Sequoia, Sequoiadendron, Metasequoia - Секвойя, секвойядендрон, метасквойя
Дендроарт
Juniperus chinensis - Можжевельник китайский, Juniperus procumbens - Можжевельник лежачий или распростертый
Wollemia - Воллемия
Microbiota - Микробиота
Sequoia, Sequoiadendron, Metasequoia - Секвойя, секвойядендрон, метасквойя
Wollemia - Воллемия
Wollemia - Воллемия
{kind=link}
